Executive Summary
Key Takeaways
- PYY(3-36) is the body's primary post-meal satiety signal, acting through Y2 receptors in the hypothalamus to suppress appetite for up to 6 hours after eating
- Individuals with obesity show 30-40% lower fasting PYY and blunted postprandial responses, contributing to impaired satiety and overeating
- Bariatric surgery (especially RYGB) increases postprandial PYY by more than 20-fold, which is a key mechanism behind sustained post-surgical weight loss
- Combined PYY and GLP-1 infusion reduces food intake by approximately 27%, exceeding the effect of either hormone alone
- Lifestyle factors including high-protein diets, dietary fiber, and aerobic exercise all increase endogenous PYY secretion
Peptide YY (PYY) is a 36-amino-acid gut hormone that functions as one of the body's most powerful natural satiety signals. Released from enteroendocrine L-cells in the distal intestine following food intake, PYY acts through the Y2 receptor in the hypothalamic arcuate nucleus to suppress appetite and reduce caloric consumption. Its role in obesity, bariatric surgery outcomes, and its potential as a therapeutic target alongside GLP-1 receptor agonists make it a central figure in modern metabolic research.
The human body produces two main forms of this peptide: PYY(1-36) and PYY(3-36). The latter is generated through enzymatic cleavage by dipeptidyl peptidase IV (DPP-IV), the same enzyme that degrades GLP-1 and GIP. PYY(3-36) is the predominant circulating form after a meal and displays selective affinity for the Y2 receptor subtype, making it the biologically active form responsible for appetite suppression. When you eat a balanced meal rich in protein and fiber, L-cells in the ileum and colon release PYY into the bloodstream within 15 to 30 minutes, with peak levels occurring approximately 1 to 2 hours postprandially and remaining elevated for up to 6 hours.
Research consistently demonstrates that individuals with obesity exhibit lower fasting PYY levels and a blunted postprandial PYY response compared to lean individuals. This deficiency creates a vicious cycle: reduced satiety signaling leads to increased caloric intake, further weight gain, and continued suppression of PYY output. The relationship between PYY and obesity is not merely correlational. Studies in PYY knockout mice show that genetic absence of this hormone leads to hyperphagia and obesity, suggesting a causative role in weight regulation. Peripheral infusion of PYY(3-36) in both lean and obese human subjects reduces food intake by approximately 30%, indicating that the PYY system remains functional even in obesity and can be therapeutically targeted.
Perhaps the most compelling evidence for PYY's importance comes from bariatric surgery research. Roux-en-Y gastric bypass (RYGB) and sleeve gastrectomy both produce dramatic increases in postprandial PYY secretion, with the area under the curve for PYY increasing more than 20-fold after RYGB. This exaggerated PYY response, occurring alongside elevated GLP-1 levels, is now considered one of the primary mechanisms behind the sustained appetite reduction and weight loss that follow bariatric surgery. The finding that pharmacological blockade of PYY and GLP-1 signaling partially reverses the appetite-suppressing effects of bypass surgery confirms the critical role these hormones play.
The combined action of PYY and GLP-1 has attracted intense pharmaceutical interest. When infused together in overweight human subjects, PYY(3-36) and GLP-1 produce additive appetite suppression greater than either hormone alone, reducing energy intake by up to 27% compared to placebo. This has prompted development of PYY analogs designed for co-administration with semaglutide and other GLP-1 receptor agonists. Novo Nordisk advanced its long-acting PYY(3-36) analog NNC0165-1875 (PYY1875) through Phase 1 and Phase 2 clinical studies, though the program was ultimately discontinued due to tolerability challenges. Nevertheless, multi-receptor agonist approaches combining GLP-1 and Y2 receptor activity remain an active area of drug design, with novel dual-agonist peptides showing promise in preclinical models.
This report provides a thorough examination of PYY biology, from its molecular structure and receptor pharmacology to its clinical significance in obesity and metabolic disease. We analyze the evidence for PYY deficiency as both a consequence and a contributing cause of obesity, review the hormonal changes that make bariatric surgery effective, and assess the current field of PYY-targeted therapeutic development. For patients and clinicians working with GLP-1 based weight management strategies, understanding PYY's complementary role provides valuable context for optimizing treatment outcomes through both pharmacological and lifestyle interventions.
Key Takeaways
- PYY(3-36) is the body's primary post-meal satiety signal, acting through Y2 receptors in the hypothalamus to suppress appetite for up to 6 hours after eating
- Individuals with obesity show 30-40% lower fasting PYY and blunted postprandial responses, contributing to impaired satiety and overeating
- Bariatric surgery (especially RYGB) increases postprandial PYY by more than 20-fold, which is a key mechanism behind sustained post-surgical weight loss
- Combined PYY and GLP-1 infusion reduces food intake by approximately 27%, exceeding the effect of either hormone alone
- Lifestyle factors including high-protein diets, dietary fiber, and aerobic exercise all increase endogenous PYY secretion
Gut Hormone Biology
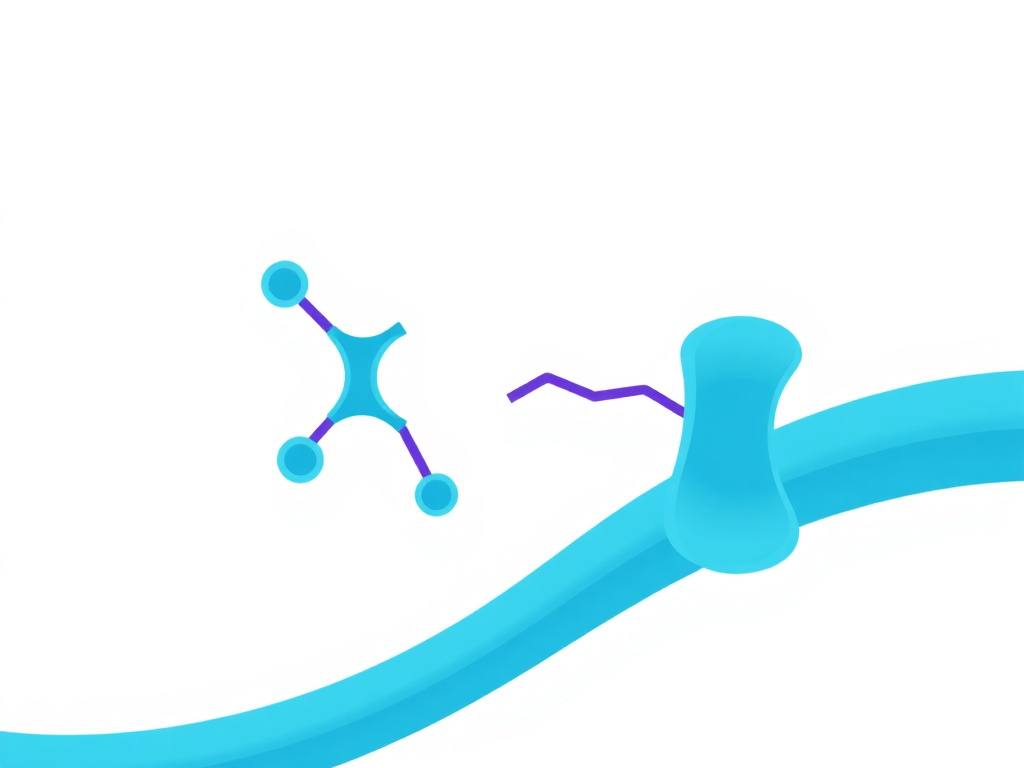
Figure 2: Enteroendocrine L-cell biology and the gut-brain signaling axis for appetite regulation
The Enteroendocrine System and L-Cell Function
The gastrointestinal tract is the largest endocrine organ in the human body, containing more than 20 distinct types of enteroendocrine cells that collectively produce over 30 hormone peptides. These cells are scattered throughout the gut epithelium from the stomach to the rectum, each type concentrated in particular regions based on its specific sensing and signaling functions. Among them, L-cells hold special importance for appetite regulation and metabolic control because they produce three key hormones: PYY, GLP-1, and GLP-2. Understanding L-cell biology is essential for anyone exploring the science behind peptide therapies.
L-cells are most densely concentrated in the distal ileum, cecum, and colon, though smaller populations exist in the duodenum and jejunum. These open-type endocrine cells have apical processes that extend into the gut lumen, allowing direct contact with ingested nutrients. When luminal nutrients reach the distal intestine, they trigger L-cell activation through multiple pathways. Glucose and amino acids stimulate L-cells through sodium-glucose cotransporter 1 (SGLT1) and various amino acid transporters, while fatty acids activate G protein-coupled receptors (GPCRs) such as GPR40 (FFAR1), GPR41 (FFAR3), GPR43 (FFAR2), and GPR120 (FFAR4) on the L-cell surface.
The signaling cascade within L-cells involves intracellular calcium elevation and cAMP generation, which trigger exocytosis of hormone-containing secretory granules. A single L-cell can co-secrete PYY, GLP-1, and GLP-2 simultaneously, though the relative proportions vary by intestinal region. Proximal L-cells tend to produce proportionally more GLP-1 relative to PYY, while distal L-cells in the colon produce higher ratios of PYY. This gradient has practical implications: nutrients that reach the distal gut more rapidly, as occurs after bariatric surgery, trigger proportionally greater PYY release.
Beyond direct nutrient sensing, L-cells also respond to neural and paracrine signals. Vagal afferent stimulation, cholinergic signaling, and bile acids all modulate L-cell secretory activity. Bile acids activate the TGR5 receptor on L-cells, explaining why bile acid sequestrants and certain bariatric procedures that alter bile acid flow affect PYY and GLP-1 secretion. Short-chain fatty acids (SCFAs) produced by bacterial fermentation of dietary fiber in the colon are potent stimulators of L-cell hormone release, binding to FFAR2 and FFAR3 receptors and increasing both PYY and GLP-1 output. This connection between gut microbiota, fiber intake, and satiety hormones has significant implications for dietary approaches to weight management.
Molecular Structure and Forms of PYY
Peptide YY belongs to the pancreatic polypeptide (PP) fold family, which also includes neuropeptide Y (NPY) and pancreatic polypeptide (PP). All three peptides share a characteristic tertiary structure known as the PP-fold, formed through specific proline and glycine residues at positions 2, 5, 8, and 9, along with tyrosine residues at positions 20 and 27. The name "peptide YY" derives from the tyrosine (Y in single-letter amino acid code) residues at both the N-terminal and C-terminal positions of the 36-amino-acid chain.
The PP-fold structure creates a hairpin-like conformation with an N-terminal polyproline helix (residues 1-8), a beta turn (residues 9-14), and an amphipathic alpha helix (residues 15-36). This three-dimensional arrangement produces a hydrophobic core that protects the peptide from rapid enzymatic degradation and is critical for receptor binding. The C-terminal alpha helix, particularly residues 25-36, contains the primary receptor-binding determinants. Mutations in this region dramatically reduce Y2 receptor affinity, while modifications to the N-terminal polyproline segment primarily affect Y1 receptor interactions.
PYY is initially synthesized as a 97-amino-acid prepropeptide in the endoplasmic reticulum of L-cells. Signal peptide cleavage yields the 67-amino-acid propeptide, which undergoes further processing by prohormone convertases (PC1/3 and PC2) to release the mature 36-amino-acid PYY(1-36). Upon secretion into the circulation, PYY(1-36) is rapidly cleaved by dipeptidyl peptidase IV (DPP-IV, also known as CD26) at the Pro2-Ile3 bond. This removes the N-terminal Tyr-Pro dipeptide to generate PYY(3-36), which accounts for approximately 60-70% of total circulating PYY immunoreactivity in the postprandial state.
The DPP-IV cleavage is not merely a degradation step but a critical activation event that alters receptor selectivity. PYY(1-36) binds with roughly equal affinity to Y1, Y2, and Y5 receptors, while PYY(3-36) displays strong selectivity for the Y2 receptor subtype, with 10-fold or greater preference over Y1 and Y5. Because Y2 receptor activation mediates appetite suppression while Y1 receptor activation can stimulate feeding, this cleavage event effectively converts PYY from a mixed agonist into a selective anorexigenic signal. Interestingly, DPP-IV inhibitors used in diabetes treatment (gliptins) can slow this conversion, potentially shifting the PYY(1-36) to PYY(3-36) ratio and affecting appetite signaling, though the clinical significance of this interaction remains debated.
The Neuropeptide Y Receptor Family
PYY exerts its biological effects through the neuropeptide Y (NPY) receptor family, a group of G protein-coupled receptors (GPCRs) that mediate the actions of NPY, PYY, and PP. In humans, four functional receptor subtypes exist: Y1, Y2, Y4, and Y5. A fifth subtype, Y6, is present in the genome but produces a truncated, non-functional protein due to a frameshift mutation. Each subtype has a distinct distribution pattern and functional role, creating a complex signaling network that regulates appetite, energy balance, and numerous other physiological processes.
The Y1 receptor is widely expressed in the central nervous system, particularly in the cerebral cortex, hippocampus, thalamus, and hypothalamus. It is also found peripherally in vascular smooth muscle, adipose tissue, and the gastrointestinal tract. Y1 activation by NPY or PYY(1-36) promotes food intake and anxiolysis. Pharmacologically, the Y1 receptor couples primarily to Gi/Go proteins, inhibiting adenylyl cyclase and reducing intracellular cAMP. It also activates phospholipase C through G-beta-gamma subunits, increasing intracellular calcium. In the context of appetite regulation, Y1 receptor activation in hypothalamic neurons stimulates feeding behavior, making it an orexigenic receptor. This is why selective Y2 binding by PYY(3-36) is crucial for appetite suppression: by activating Y2 while avoiding Y1, PYY(3-36) suppresses hunger without triggering the counterproductive feeding-stimulatory pathway.
The Y2 receptor is the primary mediator of PYY's appetite-suppressing effects. It is heavily expressed in the hypothalamic arcuate nucleus, particularly on NPY/agouti-related peptide (AgRP) neurons. Y2 receptors function as presynaptic autoreceptors on these orexigenic neurons, meaning that PYY(3-36) binding inhibits NPY release from the very neurons that drive hunger. The Y2 receptor signals through the Gi pathway, activating a beta-arrestin signaling cascade that hyperpolarizes the NPY/AgRP neuron and silences its firing. The suppression of NPY/AgRP neuronal activity disinhibits adjacent pro-opiomelanocortin (POMC) neurons, allowing increased alpha-melanocyte-stimulating hormone (alpha-MSH) release and activation of melanocortin 4 receptors (MC4R) in downstream appetite-regulating centers. This dual mechanism, direct inhibition of orexigenic signaling combined with indirect activation of anorexigenic signaling, makes Y2 receptor activation a powerful satiety signal.
The Y4 receptor shows highest affinity for pancreatic polypeptide (PP) rather than PYY, though PYY can activate it at higher concentrations. Y4 is expressed in the hypothalamus, brainstem, heart, and GI tract. Its role in appetite regulation is less well-defined than Y2, though PP-mediated Y4 activation does reduce food intake. The Y5 receptor is expressed in the hypothalamus and has been implicated in feeding stimulation, though its pharmacology is complex and context-dependent. Both NPY and PYY(1-36) activate Y5, while PYY(3-36) has lower affinity for this subtype. Some evidence suggests Y5 may play a role in long-term energy balance rather than acute meal-to-meal appetite control.
The Gut-Brain Axis: How PYY Reaches the Hypothalamus
For PYY to suppress appetite, it must communicate information from the distal gut to appetite-regulating centers in the brain. This gut-brain communication occurs through two complementary pathways: an endocrine (humoral) route and a neural (vagal) route. Understanding these pathways helps explain why different clinical interventions produce varying effects on PYY-mediated satiety.
In the endocrine pathway, PYY(3-36) released from L-cells enters the portal circulation and reaches the systemic bloodstream. Circulating PYY(3-36) crosses the blood-brain barrier through a saturable, carrier-mediated transport mechanism, gaining direct access to hypothalamic neurons in the arcuate nucleus. The arcuate nucleus occupies a unique position adjacent to the median eminence, a circumventricular organ with a partially permeable blood-brain barrier, which facilitates hormone access. Once PYY(3-36) reaches the arcuate nucleus, it binds Y2 receptors on NPY/AgRP neurons and initiates the appetite-suppressing signaling cascade described above.
The neural pathway involves vagal afferent fibers that innervate the gut wall and express Y2 receptors. PYY(3-36) released from L-cells can bind these receptors on nearby vagal nerve terminals, generating afferent signals that travel through the vagus nerve to the nucleus of the solitary tract (NTS) in the brainstem. From the NTS, the signal is relayed to the hypothalamus and other appetite-regulating brain regions including the paraventricular nucleus (PVN), the central nucleus of the amygdala (CeA), and the lateral hypothalamus. Vagotomy studies show that surgical disruption of the vagus nerve partially attenuates the appetite-suppressing effects of peripherally administered PYY(3-36), confirming the vagal contribution to PYY signaling.
A third mechanism involves PYY's effects on gastrointestinal motility. PYY activates the "ileal brake" and "colonic brake" reflexes that slow gastric emptying and upper GI transit. When you eat a large meal, PYY released from distal L-cells signals the stomach to slow emptying, allowing more complete nutrient digestion and absorption in the small intestine. This deceleration of gastric emptying contributes to prolonged feelings of fullness and reduced hunger between meals. The effect operates through both local enteric nervous system reflexes and vagal pathways, creating an integrated response that coordinates gut motility with central appetite regulation.
PYY also influences appetite through effects on reward circuitry. Functional neuroimaging studies in humans show that PYY(3-36) infusion reduces activity in the orbitofrontal cortex and striatum during food-cue presentation, suggesting decreased hedonic drive to eat. Simultaneously, PYY(3-36) increases activity in the insular cortex and hypothalamus, regions associated with interoceptive awareness and homeostatic appetite control. This shift from reward-driven eating toward homeostatic regulation may explain why PYY(3-36) reduces not only overall caloric intake but specifically high-fat and high-sugar food preference.
PYY in the Context of Gut Hormone Crosstalk
PYY does not act in isolation. It functions within an intricate network of gut hormones that collectively orchestrate appetite, digestion, and metabolic homeostasis. Understanding this network is critical for appreciating both the therapeutic potential and the complexity of targeting PYY for weight management, and it explains why combination approaches using multiple hormones, such as those explored with tirzepatide and other multi-receptor agonists, often produce superior outcomes to single-agent therapies.
GLP-1 and PYY are co-secreted from L-cells and share many regulatory functions, but they act through entirely different receptor systems. GLP-1 binds the GLP-1 receptor, a Gs-coupled GPCR that increases cAMP, while PYY(3-36) binds the Y2 receptor, a Gi-coupled GPCR that decreases cAMP. Despite these opposing intracellular signaling mechanisms, both hormones converge on appetite suppression through their respective neural targets. This convergence on a common functional outcome through distinct molecular pathways explains the additive effects observed when both hormones are administered together.
Ghrelin, often called the "hunger hormone," acts as a functional antagonist to PYY. Produced primarily by X/A-like cells in the gastric fundus, ghrelin rises before meals and falls after eating, the inverse of PYY's secretory pattern. Ghrelin activates growth hormone secretagogue receptors (GHSR) on NPY/AgRP neurons in the arcuate nucleus, stimulating the same orexigenic neurons that PYY(3-36) inhibits. The balance between ghrelin's stimulatory and PYY's inhibitory inputs to NPY/AgRP neurons is a key determinant of meal-to-meal appetite regulation. In obesity, this balance is disrupted: PYY is low while ghrelin suppression after meals is blunted, creating a pro-hunger hormonal environment.
Cholecystokinin (CCK), released from I-cells in the duodenum and jejunum, acts as an early satiety signal that promotes meal termination within 15-30 minutes of eating. CCK's rapid, short-duration action complements PYY's slower, more sustained satiety effect, creating a two-phase satiety response. Oxyntomodulin (OXM), another L-cell product, activates both GLP-1 and glucagon receptors and contributes to energy expenditure regulation. Pancreatic polypeptide (PP), released from PP cells in the pancreatic islets, preferentially activates Y4 receptors and provides additional anorexigenic input. Together, these hormones create a layered defense against overconsumption, with each hormone contributing a distinct temporal and mechanistic component to the overall satiety response.
The hormone amylin, co-secreted with insulin from pancreatic beta cells, also contributes to post-meal satiety through actions on the area postrema in the brainstem. Pramlintide, a synthetic amylin analog, has been studied in combination with GLP-1 receptor agonists for weight management. Similarly, glucagon-like peptide 2 (GLP-2), co-produced with GLP-1 and PYY in L-cells, promotes intestinal mucosal growth and nutrient absorption. While GLP-2 is not directly anorexigenic, its trophic effects on the intestinal epithelium may indirectly influence L-cell mass and hormone secretory capacity, particularly after bariatric surgery when the intestinal mucosa undergoes significant adaptive hypertrophy.
PYY & Appetite Regulation
Introduction to PYY-Mediated Appetite Control
Appetite regulation is among the most complex physiological systems in the human body, involving dozens of hormones, neuropeptides, neurotransmitters, and neural circuits that integrate information about nutritional status, energy stores, meal composition, and environmental context. Within this system, PYY occupies a position of particular importance as a post-meal satiety signal that bridges the gut and the brain. Its discovery as an appetite regulator transformed our understanding of how the body controls food intake, shifting focus from purely mechanical stomach-stretching models to hormonal signaling frameworks that continue to drive therapeutic innovation.
The significance of PYY in appetite regulation extends beyond its direct effects on food intake. PYY influences meal duration, inter-meal intervals, food preference, portion size, and even the conscious experience of hunger and satiety. It modulates not only homeostatic appetite (eating driven by caloric need) but also hedonic appetite (eating driven by pleasure and reward). This breadth of action makes PYY relevant to nearly every clinical scenario involving disordered eating behavior, from the hyperphagia of genetic obesity syndromes to the emotional eating patterns common in mild-to-moderate obesity. For clinicians and patients working with GLP-1 based treatments, understanding PYY's complementary role provides a more complete picture of the appetite regulation system being targeted.
This section examines the evidence for PYY's role in appetite regulation from its initial discovery through current understanding, covering central nervous system mechanisms, temporal dynamics, reward system interactions, dose-response relationships, and interactions with other appetite-regulating hormones. Each topic has direct relevance to clinical weight management and helps explain why certain dietary, exercise, and pharmacological interventions succeed or fail in controlling appetite.
The Discovery of PYY's Anorexigenic Properties
Peptide YY was first isolated from porcine intestinal extracts in 1980 by Tatemoto and Mutt, who identified it based on its C-terminal tyrosine amide residue. However, its role in appetite regulation was not recognized for more than two decades. The seminal 2002 study by Batterham and colleagues at University College London fundamentally changed understanding of PYY's physiological function. They demonstrated that peripheral administration of PYY(3-36) to both rodents and humans produced significant reductions in food intake, establishing PYY as a key endogenous satiety hormone. In their human study, intravenous infusion of PYY(3-36) to physiological postprandial levels reduced caloric intake at a buffet meal by 36% in lean volunteers. This finding, published in Nature, sparked intense research interest in PYY as both a regulator of appetite and a potential therapeutic target.
The initial excitement was tempered somewhat when several groups reported difficulty replicating the appetite-suppressing effects of PYY(3-36) in rodent models, particularly when using non-stressed animals in familiar feeding environments. This controversy highlighted the importance of experimental conditions and stress responses in appetite research. However, subsequent carefully controlled studies confirmed PYY's anorexigenic effects across species, and the consensus now firmly supports PYY(3-36) as a physiologically relevant satiety signal. The key qualification is that PYY's effects are dose-dependent and can be influenced by psychological state, baseline energy balance, and the macronutrient composition of recent meals.
Central Mechanisms of Appetite Suppression
PYY(3-36) suppresses appetite through a well-characterized central nervous system pathway that begins in the hypothalamic arcuate nucleus and propagates through several downstream brain regions. The arcuate nucleus contains two functionally opposed neuronal populations: the orexigenic NPY/AgRP neurons and the anorexigenic POMC neurons. PYY(3-36) targets the Y2 receptors expressed on NPY/AgRP neurons, which function as presynaptic inhibitory autoreceptors.
When PYY(3-36) binds to Y2 receptors on NPY/AgRP neurons, it activates the Gi/Go signaling pathway. This reduces intracellular cAMP levels, inhibits voltage-gated calcium channels (primarily N-type and P/Q-type), and activates G protein-coupled inwardly rectifying potassium (GIRK) channels. The net effect is membrane hyperpolarization and reduced action potential firing. The silenced NPY/AgRP neuron releases less NPY (which would otherwise stimulate feeding via Y1 and Y5 receptors in downstream targets), less AgRP (which would otherwise block MC4R-mediated satiety signaling), and less GABA (which provides tonic inhibition of neighboring POMC neurons).
The reduction in GABAergic tone from NPY/AgRP neurons disinhibits POMC neurons, allowing increased transcription and processing of the POMC precursor polypeptide. POMC processing generates alpha-MSH, which is released onto melanocortin 4 receptors (MC4R) in the paraventricular nucleus (PVN) and other hypothalamic regions. MC4R activation initiates a strong anorexigenic signal that reduces meal size, increases inter-meal intervals, and promotes energy expenditure. Genetic mutations in MC4R cause monogenic obesity in humans, underscoring the critical importance of this pathway. PYY's ability to engage this system indirectly, through NPY/AgRP neuron inhibition, makes it an important upstream regulator of melanocortin signaling.
Beyond the hypothalamus, PYY(3-36) influences appetite through brainstem circuits. The nucleus of the solitary tract (NTS) and area postrema receive vagal afferent input carrying PYY signals from the gut, as well as direct humoral PYY exposure due to the area postrema's location outside the blood-brain barrier. These brainstem nuclei process visceral information about gut distension, nutrient content, and hormonal status, integrating this input with hypothalamic signals to generate a coordinated appetite response. The NTS projects to the parabrachial nucleus (PBN), which in turn communicates with the central nucleus of the amygdala (CeA) and the bed nucleus of the stria terminalis (BNST), brain regions involved in aversive and emotional responses to food intake and satiety.
Temporal Dynamics of PYY Release and Satiety
The pattern of PYY secretion following a meal follows a characteristic biphasic profile that has important implications for both physiological satiety and therapeutic applications. An initial rapid phase begins within 15-30 minutes of eating, before ingested nutrients have reached the distal gut where most L-cells reside. This early release is mediated by neural reflexes, particularly vagal signaling from the proximal gut, and by paracrine signaling from more proximally located L-cells. Proximal L-cells in the duodenum and jejunum, while fewer in number, can respond rapidly to arriving nutrients and initiate the early PYY response.
The second, larger phase of PYY release occurs 1-2 hours after meal ingestion, coinciding with nutrient arrival in the ileum and colon where L-cell density is highest. This sustained release maintains elevated PYY levels for 4-6 hours, providing prolonged satiety signaling that extends well beyond the period of active eating. The magnitude and duration of the PYY response correlate with several meal characteristics. Caloric load is the primary determinant: larger meals produce proportionally greater PYY secretion. Macronutrient composition also matters. Protein produces the strongest PYY response per calorie, followed by fat, then carbohydrate. High-fiber meals produce a delayed but prolonged PYY response due to colonic fermentation of fiber to short-chain fatty acids, which continue to stimulate L-cells for hours after the meal.
The practical implications are clear for anyone managing appetite, whether through dietary strategies alone or in combination with semaglutide or other GLP-1 therapies. Meals structured around protein and fiber will naturally amplify PYY release and prolong satiety. This aligns with the clinical observation that high-protein diets produce greater weight loss and better appetite control than isocaloric low-protein diets, an effect partially mediated by enhanced PYY secretion.
PYY and the Hedonic System
Appetite regulation involves not only homeostatic mechanisms (hunger/satiety) but also hedonic (reward-based) mechanisms that drive eating for pleasure rather than caloric need. PYY influences both systems, which is critical for its potential therapeutic value. Functional magnetic resonance imaging (fMRI) studies have revealed that PYY(3-36) infusion alters brain activation patterns in response to food cues. In fed subjects receiving PYY(3-36), activity decreases in the orbitofrontal cortex (OFC), striatum, and ventral tegmental area (VTA), regions comprising the mesolimbic dopamine reward pathway. These are the same circuits activated by palatable food images and flavors, and their suppression by PYY correlates with reduced desire to eat and decreased food appeal ratings.
Simultaneously, PYY(3-36) increases activation in the posterior insula and hypothalamus, regions associated with interoceptive awareness and homeostatic appetite control. This shift, away from reward-driven eating and toward physiological satiety, may explain clinical observations that PYY(3-36) reduces preference for high-fat, energy-dense foods more than for bland or low-calorie options. If PYY only affected homeostatic hunger, you would expect proportional reductions across all food types. The selective reduction in hedonic eating suggests PYY(3-36) modulates the motivational value of food at a cortical level.
This dual action on homeostatic and hedonic circuits distinguishes PYY from some other satiety signals. CCK, for example, primarily affects meal termination through vagal brainstem circuits without significant hedonic modulation. GLP-1 receptor agonists like liraglutide and semaglutide also engage both homeostatic and hedonic systems, acting on GLP-1 receptors in both hypothalamic and reward-processing brain regions. The convergent but mechanistically distinct effects of PYY and GLP-1 on reward circuitry provide a rationale for combination therapy: by engaging reward suppression through two different receptor systems (Y2 and GLP-1R), the combined effect may be more complete and harder to overcome by compensatory mechanisms.
Dose-Response Relationships and Pharmacology
Human infusion studies have established clear dose-response relationships for PYY(3-36)'s appetite-suppressing effects. At physiological postprandial concentrations (approximately 30-50 pmol/L), PYY(3-36) produces moderate appetite suppression equivalent to a small meal. At supraphysiological levels (80-100 pmol/L), the effect is more pronounced, with food intake reductions of 30-36% observed in multiple controlled studies. However, doses producing plasma levels above approximately 100-150 pmol/L begin to cause nausea and malaise, limiting the therapeutic window.
The nausea associated with high-dose PYY(3-36) is believed to be mediated by area postrema activation, similar to the mechanism underlying GLP-1 receptor agonist-associated nausea. This overlap in side effect profiles has proven to be a significant challenge for combination therapy development. When PYY(3-36) analogs are co-administered with semaglutide, the gastrointestinal side effects appear to be at least partially additive, narrowing the window between effective appetite suppression and intolerable nausea. This was a key factor in Novo Nordisk's decision to discontinue clinical development of PYY1875 as an add-on to semaglutide.
The half-life of native PYY(3-36) in circulation is relatively short, approximately 7-15 minutes, due to renal clearance and further enzymatic degradation by aminopeptidases and endopeptidases. This rapid clearance has driven the development of long-acting PYY analogs with modifications to resist enzymatic degradation and reduce renal filtration. Strategies include fatty acid acylation (to promote albumin binding and extend half-life), PEGylation, amino acid substitution at cleavage-susceptible positions, and fusion with Fc antibody fragments. These modifications have achieved half-lives ranging from hours to days in preclinical and early clinical studies, though maintaining receptor selectivity and avoiding off-target effects remains an engineering challenge.
PYY Interactions with Other Appetite Hormones
PYY's appetite-suppressing effects are modulated by the concurrent hormonal milieu. Ghrelin, the primary orexigenic gut hormone, functionally antagonizes PYY at the level of the arcuate nucleus. When ghrelin levels are high (as in the pre-meal fasted state), the strong activation of NPY/AgRP neurons partially overcomes PYY-mediated inhibition, even if PYY levels are moderately elevated. This dynamic interplay means that PYY is most effective at suppressing appetite when ghrelin levels have been appropriately suppressed, as occurs after a substantial meal. It also explains why PYY infusion is less effective at reducing food intake in profoundly fasted individuals: the strong orexigenic drive from high ghrelin levels competes with PYY's anorexigenic signal.
GLP-1 and PYY co-secreted from L-cells amplify each other's effects on appetite through convergent but non-overlapping neural circuits. Studies using specific receptor antagonists have shown that blocking either GLP-1 receptors or Y2 receptors partially attenuates the satiety response to a meal, but blocking both abolishes it nearly completely. This indicates that the two hormones contribute independently and additively to post-meal satiety, a finding with direct implications for GLP-1 receptor agonist therapy. Patients on GLP-1 receptor agonists who also optimize their endogenous PYY release through dietary strategies (high protein, high fiber, regular meals) may experience enhanced satiety beyond what the drug alone provides.
Insulin, leptin, and adiponectin provide longer-term adiposity signals that modulate the sensitivity of hypothalamic neurons to acute meal-related hormones including PYY. Chronic leptin signaling downregulates NPY expression in arcuate neurons, which can alter the baseline against which PYY's inhibitory effects operate. In leptin-resistant obesity, NPY/AgRP neurons are tonically hyperactive, and higher concentrations of PYY(3-36) may be needed to achieve the same degree of appetite suppression seen in lean individuals. This concept of "PYY resistance" has been proposed by analogy with leptin and insulin resistance, though the evidence is less definitive. What is clear is that obese individuals require higher absolute levels of PYY(3-36) infusion to achieve the same subjective appetite suppression as lean controls, consistent with some degree of reduced sensitivity.
Amylin, co-secreted with insulin from pancreatic beta cells, engages the area postrema to reduce meal size and slow gastric emptying. Its mechanism partially overlaps with PYY's brainstem-mediated effects, and combined administration of amylin with PYY(3-36) in preclinical models produces additive appetite suppression. Cagrilintide, a long-acting amylin analog being developed by Novo Nordisk, combined with semaglutide (CagriSema) has produced impressive weight loss results in clinical trials. The success of amylin-GLP-1 combinations provides proof of concept for multi-hormone approaches to weight management, supporting continued interest in PYY as an additional component of future combination therapies.
PYY and Gastrointestinal Motility
Beyond its central appetite-suppressing actions, PYY exerts profound effects on gastrointestinal motility that contribute to satiety through peripheral mechanisms. PYY activates what physiologists term the "ileal brake" and "colonic brake," reflexes that slow upper GI transit in response to nutrient detection in the distal intestine. When PYY is released from L-cells in the ileum and colon, it signals the stomach and proximal small intestine to decelerate their motor activity, reducing gastric emptying rate, decreasing small intestinal peristalsis, and increasing the time nutrients spend in contact with absorptive surfaces.
The ileal brake operates through both neural and hormonal pathways. PYY acts on enteric nervous system neurons within the gut wall, modulating the peristaltic reflex through inhibitory Y2 receptor-expressing interneurons. It also activates vagal afferent fibers that transmit inhibitory signals to the dorsal motor nucleus of the vagus, reducing vagal efferent drive to the stomach. The combined effect is a measurable slowing of gastric emptying that can be quantified using scintigraphic or breath test methods. Studies show that PYY(3-36) infusion at physiological doses delays gastric emptying half-time by 15-30 minutes, a deceleration that has meaningful effects on postprandial glucose excursions, nutrient absorption kinetics, and subjective fullness.
The colonic brake mechanism is less well characterized but involves PYY-mediated reduction in colonic motility and secretion. PYY reduces colonic propulsive activity and decreases water and electrolyte secretion, which concentrates colonic contents and slows transit. These effects, combined with PYY's central appetite-suppressing actions, create an integrated physiological response: the brain receives signals to stop eating while the gut simultaneously adjusts its processing speed to match the volume and composition of nutrients already consumed. This coordination ensures that satiety signaling aligns with actual digestive capacity, preventing both overconsumption and digestive overwhelm.
For patients using semaglutide or other GLP-1 receptor agonists, the GI motility effects of PYY are particularly relevant because GLP-1 receptor agonists also slow gastric emptying. The combination of drug-induced and PYY-mediated gastric emptying delay can sometimes produce excessive upper GI stasis, manifesting as persistent nausea, bloating, early satiety beyond therapeutic intent, or rarely gastroparesis. Understanding that both PYY and GLP-1 contribute to gastric emptying regulation helps clinicians anticipate and manage these effects, for example by adjusting meal composition (smaller, more frequent, lower-fat meals) to reduce the total motility-slowing stimulus during the early weeks of GLP-1 receptor agonist therapy when PYY-mediated responses are still recalibrating.
PYY in Intestinal Immunity and Barrier Function
Emerging research has identified roles for PYY beyond metabolism and appetite, particularly in intestinal immune regulation and barrier function. Y1 and Y2 receptors are expressed on various immune cell populations in the gut-associated lymphoid tissue (GALT), including dendritic cells, macrophages, and T lymphocytes. PYY modulates immune cell trafficking, cytokine production, and mucosal immune responses, suggesting that PYY deficiency in obesity may contribute to the chronic low-grade intestinal inflammation observed in metabolic disease.
PYY also influences intestinal epithelial barrier integrity. Studies in rodent models show that PYY promotes tight junction protein expression and reduces intestinal permeability. This is relevant to the "leaky gut" hypothesis of obesity-related metabolic inflammation, which proposes that increased intestinal permeability allows bacterial endotoxins (lipopolysaccharide, LPS) to enter the systemic circulation, triggering chronic inflammation that contributes to insulin resistance and metabolic dysfunction. By maintaining barrier integrity, adequate PYY levels may protect against this pathogenic cascade. The restoration of PYY levels after bariatric surgery could therefore contribute to the anti-inflammatory effects of surgery through improved intestinal barrier function, in addition to the well-characterized appetite and metabolic benefits.
These non-metabolic functions of PYY highlight the breadth of L-cell hormones' physiological roles and provide additional rationale for strategies that optimize PYY secretion through dietary fiber, which also supports microbiome health and barrier function. The biohacking hub explores these connections between gut health, peptide hormones, and overall metabolic wellness.
Circadian Rhythms and PYY Secretion
PYY secretion follows a circadian pattern that interacts with meal timing to produce time-of-day variations in appetite and satiety. Basal PYY levels are lowest in the early morning, rise modestly through the morning hours even in the absence of food intake, and reach their highest baseline levels in the late afternoon and evening. This circadian variation is superimposed on the meal-driven PYY responses, creating a complex temporal pattern that influences when people feel most and least hungry.
The circadian PYY rhythm has practical implications for meal timing strategies. The lower baseline PYY in the morning means that breakfast may need to be particularly protein-rich and fiber-rich to generate a sufficient PYY response and suppress mid-morning hunger effectively. The higher baseline PYY in the evening contributes to the observation that people often find it easier to eat moderate evening meals than moderate breakfasts, though this effect is counterbalanced by increased hedonic drive and stress-related eating that typically peak in the evening hours.
Sleep deprivation disrupts the circadian PYY rhythm, reducing the amplitude of the evening peak and lowering overall 24-hour PYY exposure. This disruption contributes to the increased hunger, decreased satiety, and preference for high-calorie foods consistently observed in sleep-deprived individuals. A single night of total sleep deprivation can reduce postprandial PYY levels by 15-20% the following day, increasing ad libitum food intake by 200-500 calories. Chronic partial sleep restriction (sleeping 5-6 hours per night versus 7-9 hours) produces sustained reductions in PYY and corresponding increases in daily caloric intake, contributing to weight gain over time. These findings emphasize the importance of adequate sleep as a component of any weight management strategy, whether pharmacological or lifestyle-based.
Shift work represents a particularly challenging scenario for PYY-mediated appetite regulation. Workers on rotating shifts eat at times misaligned with their circadian PYY rhythms, potentially experiencing reduced postprandial PYY responses to meals consumed during the biological night. This circadian misalignment may contribute to the increased obesity risk well-documented in shift workers. Strategies to mitigate this effect include emphasizing protein-rich meals during biological night shifts and maintaining consistent meal timing within each shift rotation to establish secondary entrained PYY rhythms.
Obesity & PYY Deficiency
PYY-Mediated Satiety Response
Evidence for Reduced PYY in Obesity
One of the most consistent findings in obesity research is that individuals with elevated body mass index (BMI) display significantly lower circulating PYY levels compared to lean counterparts. This reduction affects both fasting (baseline) PYY concentrations and the postprandial PYY response to meals. Multiple cross-sectional studies spanning different populations, age groups, and ethnic backgrounds have confirmed this association, establishing PYY deficiency as a reliable biomarker of the obese state.
In quantitative terms, fasting PYY levels in individuals with BMI greater than 30 kg/m2 are typically 20-40% lower than those in normal-weight individuals (BMI 18.5-24.9 kg/m2). The postprandial rise in PYY after a standardized meal is even more dramatically reduced, with obese individuals showing a blunted peak response that is 40-60% lower than lean controls. The area under the curve (AUC) for postprandial PYY, a measure of total PYY exposure over the hours following a meal, is correspondingly reduced. This means obese individuals experience less total satiety signaling from PYY during and after eating, potentially contributing to larger meal sizes, shorter inter-meal intervals, and greater daily caloric intake.
The relationship between BMI and PYY is approximately linear across the weight spectrum. Even overweight individuals (BMI 25-29.9) show moderately reduced PYY compared to those at normal weight, and morbidly obese individuals (BMI > 40) display the most pronounced deficits. Waist circumference, a marker of visceral adiposity, shows an even stronger inverse correlation with PYY levels than BMI alone, suggesting that central fat distribution is particularly associated with impaired PYY secretion. This aligns with the known metabolic toxicity of visceral adipose tissue and its association with insulin resistance, inflammation, and other hormonal disruptions.
Cause or Consequence: The Chicken-or-Egg Question
A fundamental question in PYY research is whether reduced PYY levels in obesity represent a cause of weight gain, a consequence of it, or both. Several lines of evidence inform this debate, and the current consensus favors a bidirectional relationship in which PYY deficiency both contributes to and is worsened by obesity.
Evidence supporting PYY deficiency as a contributing cause of obesity comes from several sources. First, PYY knockout mice develop hyperphagia and obesity on a standard chow diet, demonstrating that genetic absence of PYY is sufficient to produce excessive weight gain even without caloric excess. Second, transgenic mice overexpressing PYY are resistant to diet-induced obesity, maintaining normal body weight even when provided unlimited access to high-fat diets. Third, a study in insulin-sensitive, non-diabetic subjects with a strong family history of type 2 diabetes found blunted postprandial PYY responses despite normal body weight and insulin sensitivity. This suggests that PYY deficiency can precede the development of obesity and metabolic disease in genetically susceptible individuals, pointing toward a primary rather than secondary role.
However, evidence also supports PYY reduction as a consequence of obesity. Diet-induced obese rodents develop reduced PYY secretion over time as obesity progresses, and weight loss through caloric restriction partially restores PYY levels. In humans, weight loss following bariatric surgery produces dramatic increases in PYY secretion (discussed in detail in the bariatric surgery section), though this is partly due to anatomical changes that alter nutrient delivery to L-cells. The relationship with insulin resistance is particularly revealing: low serum PYY has been linked to insulin resistance in first-degree relatives of subjects with type 2 diabetes, and PYY(AUC) correlates positively with insulin sensitivity. Since insulin resistance is both a driver and a consequence of obesity, PYY deficiency may participate in the same feed-forward cycle.
The most likely model is a positive feedback loop: genetic or early-life factors produce relatively lower PYY secretory capacity in certain individuals, predisposing them to increased caloric intake and weight gain. As obesity develops, chronic metabolic changes including insulin resistance, low-grade inflammation, and altered gut microbiome composition further suppress PYY secretion, deepening the deficit and accelerating weight gain. This vicious cycle explains why obesity is self-reinforcing and why breaking the cycle, whether through surgery, pharmacotherapy with agents like semaglutide, or intensive lifestyle intervention, often produces improvements in PYY levels alongside weight loss.
PYY, Insulin Resistance, and Type 2 Diabetes
The relationship between PYY and insulin resistance extends beyond shared associations with obesity. PYY directly modulates insulin secretion and peripheral insulin sensitivity, creating a metabolic link between appetite regulation and glucose homeostasis. PYY suppresses insulin secretion from pancreatic beta cells through Y1 receptor activation on islet cells, while simultaneously enhancing peripheral tissue insulin sensitivity. This dual action produces a phenotype of lower insulin levels with preserved glucose disposal, the metabolic hallmark of insulin-sensitive lean physiology.
In type 2 diabetes, PYY levels are typically even lower than in obesity without diabetes, and the postprandial PYY response is severely blunted. Studies comparing obese individuals with and without type 2 diabetes show that diabetes confers an additional PYY deficit beyond what is explained by BMI alone. Plasma PYY levels after consuming a mixed meal are significantly reduced in subjects with type 2 diabetes compared to non-diabetic controls matched for body weight. This suggests that the metabolic derangements specific to diabetes, including chronic hyperglycemia, hyperinsulinemia, and altered incretin signaling, further impair L-cell PYY secretion.
The practical relevance for patients on GLP-1 receptor agonist therapy is substantial. Tirzepatide, a dual GIP/GLP-1 receptor agonist, has shown the most impressive weight loss results of any anti-obesity medication tested to date, with mean weight loss of 20-25% in the SURMOUNT trials. Part of tirzepatide's efficacy may relate to its effects on the broader gut hormone milieu. GLP-1 receptor agonists slow gastric emptying and alter nutrient delivery to the distal gut, potentially influencing endogenous PYY secretion patterns. Whether GLP-1 receptor agonists increase, decrease, or have neutral effects on PYY secretion is still debated, with study results varying based on the specific drug, dose, and duration of treatment.
PYY Deficiency Across Special Populations
The pattern of PYY deficiency in obesity extends to several special populations that merit individual discussion. Prader-Willi syndrome (PWS), a genetic disorder characterized by extreme hyperphagia and obesity from early childhood, is associated with profoundly reduced fasting PYY and a blunted postprandial PYY response. Individuals with PWS show a lesser suppression of pre-meal ghrelin alongside their reduced PYY, creating a doubly adverse hormonal profile: excess hunger signaling combined with insufficient satiety signaling. This hormonal imbalance contributes to the insatiable appetite that makes PWS among the most challenging obesity conditions to manage.
In polycystic ovary syndrome (PCOS), which is frequently associated with obesity and insulin resistance, PYY levels are reduced compared to weight-matched controls without PCOS. The hyperandrogenism and insulin resistance characteristic of PCOS may contribute to impaired L-cell function, though the mechanisms are not fully elucidated. Women with PCOS who achieve weight loss through any means, including lifestyle modification, metformin, or GLP-1 receptor agonists, typically show improvements in PYY levels alongside reductions in androgens and improvements in insulin sensitivity.
Elderly individuals generally show declining PYY levels with age, though paradoxically some studies report that the anorexigenic effect of a given PYY level is enhanced in older adults. This increased sensitivity may contribute to the "anorexia of aging" seen in some elderly populations. In pediatric obesity, the PYY deficit mirrors that seen in adults, with obese children showing lower fasting and postprandial PYY compared to normal-weight peers. Early intervention to restore normal PYY dynamics through dietary modification (increased protein and fiber, reduced refined carbohydrate) may be particularly valuable in pediatric populations, potentially interrupting the obesity-perpetuating feedback loop before it becomes entrenched.
Dietary and Lifestyle Factors Affecting PYY Levels
One of the most actionable aspects of PYY research is the identification of dietary and lifestyle factors that enhance endogenous PYY secretion. For individuals using pharmacological weight management strategies, optimizing these natural PYY-boosting approaches can provide additive benefits and improve treatment outcomes. Anyone working with a weight management program should consider these evidence-based strategies.
Dietary protein is the most potent macronutrient stimulus for PYY secretion. Isocaloric meals with 30-40% protein produce approximately 30-50% higher postprandial PYY levels compared to meals with 10-15% protein. The mechanism involves both direct amino acid sensing by L-cells and indirect effects through altered gastric emptying and small intestinal transit. Among protein sources, whey protein and casein appear to produce the strongest PYY responses, though the differences between protein types are smaller than the differences between high-protein and low-protein meals overall. From a practical standpoint, including 25-40 grams of protein per meal is sufficient to meaningfully enhance PYY release.
Dietary fiber, particularly soluble and fermentable fiber, stimulates PYY through multiple mechanisms. Viscous soluble fibers slow gastric emptying and enhance nutrient contact with L-cells in the small intestine. Fermentable fibers (inulin, fructooligosaccharides, resistant starch, beta-glucan) are metabolized by colonic bacteria to produce short-chain fatty acids, primarily acetate, propionate, and butyrate. These SCFAs bind FFAR2 and FFAR3 receptors on colonic L-cells, triggering sustained PYY secretion that can persist for hours after the meal. A daily fiber intake of 25-35 grams from diverse sources provides optimal stimulation of both small intestinal and colonic L-cells.
Physical exercise acutely increases PYY levels, with aerobic exercise showing more consistent effects than resistance training. High-intensity interval training (HIIT) produces the strongest PYY elevations, with levels rising during and immediately after exercise and remaining elevated for 1-3 hours post-exercise. This exercise-induced PYY rise partially explains the transient appetite suppression commonly reported after vigorous physical activity, sometimes called "exercise-induced anorexia." Regular aerobic exercise training appears to enhance the postprandial PYY response over time, suggesting adaptive improvements in L-cell sensitivity or secretory capacity with chronic training.
Meal timing and frequency also influence PYY dynamics. Regular meal patterns with consistent timing produce more predictable and potentially more effective PYY responses than erratic eating schedules. Intermittent fasting approaches show variable effects on PYY: the prolonged fast produces very low PYY levels, but the refeeding meal after a fast typically generates a strong PYY surge. Whether this compensatory surge is sufficient to offset the low-PYY fasting period in terms of overall appetite control varies between individuals and fasting protocols.
Sleep quality and duration affect PYY through circadian and metabolic pathways. Sleep deprivation reduces postprandial PYY levels and increases hunger, contributing to the well-documented association between poor sleep and weight gain. Ensuring 7-9 hours of quality sleep supports normal PYY circadian rhythms and maximizes the satiety response to meals. Stress, through cortisol elevation and sympathetic activation, can also impair PYY secretion and enhance ghrelin release, creating a hormonal environment that promotes overeating. Stress management strategies including mindfulness, adequate sleep, and regular physical activity therefore have indirect benefits for PYY-mediated appetite regulation.
PYY and the Gut Microbiome Connection in Obesity
The gut microbiome plays a significant role in modulating PYY levels, and the altered microbiome composition (dysbiosis) observed in obesity may contribute to PYY deficiency through reduced short-chain fatty acid (SCFA) production. In lean individuals with a diverse, fiber-rich diet, colonic bacteria produce substantial quantities of acetate, propionate, and butyrate from the fermentation of dietary fiber and resistant starch. These SCFAs activate free fatty acid receptors (FFAR2 and FFAR3) on colonic L-cells, stimulating PYY and GLP-1 release. Propionate, in particular, has been shown to be a potent PYY secretagogue in both rodent and human studies.
Obesity is associated with reduced microbial diversity and altered phylum-level composition, typically characterized by a decreased Bacteroidetes-to-Firmicutes ratio (though this specific ratio has been debated in more recent studies). More critically, obesity-associated dysbiosis is often characterized by reduced abundance of SCFA-producing bacterial species, including Faecalibacterium prausnitzii, Roseburia intestinalis, and Eubacterium rectale. The resulting decrease in colonic SCFA production may directly reduce L-cell PYY output, contributing to the overall PYY deficiency seen in obesity.
Interventions that restore microbial SCFA production capacity can modestly increase PYY levels. Prebiotic supplementation with inulin (10-15 grams daily) has been shown to increase both fasting and postprandial PYY in overweight adults, with corresponding improvements in subjective appetite ratings and small but statistically significant reductions in caloric intake. Probiotic supplementation with specific Lactobacillus and Bifidobacterium strains has produced more variable results, with some studies showing PYY increases and others showing no effect. The variability likely reflects strain-specific differences in SCFA production capacity and the influence of baseline microbiome composition on probiotic colonization success.
Fecal microbiota transplantation (FMT) from lean donors to obese recipients has been investigated for metabolic effects, with some studies reporting improvements in insulin sensitivity and modest changes in gut hormone profiles including PYY. However, the effects are generally small and transient, suggesting that sustained microbiome modification requires ongoing dietary support (fiber intake) to maintain the transplanted community. The connection between microbiome, SCFAs, and PYY provides a compelling rationale for emphasizing dietary fiber as a cornerstone of weight management, whether used alone or in combination with pharmacotherapy like semaglutide.
The Role of Inflammation in PYY Suppression
Chronic low-grade inflammation, a hallmark of obesity, may directly impair L-cell PYY secretion through several mechanisms. Elevated circulating levels of pro-inflammatory cytokines including tumor necrosis factor alpha (TNF-alpha), interleukin-6 (IL-6), and interleukin-1 beta (IL-1 beta) have been shown to reduce PYY gene expression and secretion in cell culture and animal models. These cytokines activate NF-kB and STAT3 signaling pathways in L-cells, suppressing the transcription of the PYY gene and reducing the number of PYY-containing secretory granules available for release.
Adipose tissue inflammation, particularly in visceral fat depots, is a major source of circulating pro-inflammatory cytokines in obesity. The inverse correlation between waist circumference and PYY levels, which is stronger than the correlation between BMI and PYY, may partly reflect this mechanism: visceral adiposity produces more inflammatory signaling than subcutaneous adiposity, leading to greater suppression of L-cell PYY output. Weight loss, through any mechanism, reduces adipose tissue inflammation and may thereby relieve inflammatory suppression of PYY secretion, contributing to the improved appetite regulation often experienced after significant weight reduction.
This inflammatory mechanism also helps explain why metabolic surgery produces such rapid improvements in PYY signaling. Beyond the anatomical changes that increase nutrient delivery to L-cells, bariatric surgery rapidly reduces systemic inflammation, with measurable decreases in CRP, IL-6, and TNF-alpha within days of the procedure, before significant weight loss has occurred. This anti-inflammatory effect may "unblock" L-cell PYY production, contributing to the very early appetite changes reported by post-surgical patients. Anti-inflammatory dietary patterns (Mediterranean diet, omega-3 fatty acid supplementation) may provide a milder version of this effect, supporting PYY secretion through reduced inflammatory suppression. The lifestyle hub provides guidance on anti-inflammatory eating approaches that complement weight management strategies.
Genetic Factors in PYY Production and Response
Genetic variation influences both PYY secretory capacity and responsiveness to PYY signaling, contributing to individual differences in appetite regulation and obesity susceptibility. Several single nucleotide polymorphisms (SNPs) in the PYY gene and its regulatory regions have been associated with differences in circulating PYY levels, body weight, and eating behavior. A polymorphism in the PYY promoter region affects transcription factor binding and PYY gene expression, with the minor allele associated with lower PYY levels and increased obesity risk in some populations.
Variants in the NPY2R gene (encoding the Y2 receptor) affect receptor expression, signaling efficiency, and downstream appetite responses to PYY. Individuals carrying specific NPY2R variants show reduced anorexigenic responses to PYY(3-36) infusion despite achieving similar plasma drug levels, suggesting receptor-level differences in PYY sensitivity. These genetic variants may partially explain why some individuals are more responsive than others to dietary and lifestyle interventions that boost PYY, and why the weight loss response to bariatric surgery, which relies heavily on enhanced PYY signaling, varies substantially between patients.
Epigenetic modifications to the PYY gene have been identified in studies of maternal nutrition and offspring metabolic programming. In animal models, maternal high-fat diet during pregnancy and lactation produces epigenetic changes (DNA methylation, histone modification) at the PYY locus in offspring, reducing PYY expression and increasing obesity susceptibility in adulthood. These findings suggest that some cases of low PYY and obesity may have developmental origins, programmed in utero or early postnatal life by maternal metabolic conditions. While these epigenetic changes are potentially modifiable through environmental interventions, they represent a biological predisposition that may require more intensive therapeutic approaches to overcome.
Understanding the genetic and epigenetic field of PYY biology has implications for precision medicine approaches to weight management. In the future, genetic profiling of PYY-related variants could help predict which patients would benefit most from PYY-augmenting strategies (dietary fiber, exercise, microbiome modulation) versus those who might require pharmacological appetite suppression through different pathways. Current assessment tools can help identify individual factors that influence treatment selection, and genetic considerations may eventually be incorporated into these assessments.
GLP-1 + PYY Combined Effects

Evidence for Additive Appetite Suppression
The observation that PYY and GLP-1 are co-secreted from the same L-cells suggested early on that they might function as a coordinated appetite-suppressing system. Testing this hypothesis required controlled infusion studies in which each hormone was administered alone and in combination, allowing quantification of individual and combined effects. The results have consistently demonstrated that PYY(3-36) and GLP-1 produce additive appetite suppression when administered together, sometimes exceeding what would be predicted from simple addition of their individual effects.
The landmark study by Neary and colleagues (2005) infused PYY(3-36) and GLP-1(7-36 amide) individually and in combination to healthy overweight men. Neither PYY(3-36) alone nor GLP-1 alone at the doses used significantly reduced energy intake compared to saline placebo. However, combined infusion of both hormones at these same individually sub-threshold doses reduced energy intake by approximately 27% compared to placebo. This was a critical finding because it demonstrated that the two hormones cooperate at doses below their individual effective thresholds, suggesting true physiological cooperation rather than merely additive pharmacological effects. The combined infusion also produced greater subjective ratings of fullness and reduced ratings of hunger and prospective food consumption compared to either hormone alone.
Subsequent studies have refined these findings. Schmidt and colleagues (2014) showed that combined PYY(3-36) and GLP-1 infusion reduced ad libitum energy intake, with the combination producing effects that were additive. They also demonstrated that the combined infusion reduced appetite-related neural activation in fMRI studies, with greater suppression of activity in the insula and orbitofrontal cortex than either hormone alone. These brain regions are involved in processing food-related reward and interoceptive signals, indicating that the combined hormonal signal produces a qualitatively stronger satiety response at the neural level.
Distinct Receptor Mechanisms Enabling Combined Effects
The additive effects of PYY and GLP-1 arise from their activation of entirely separate receptor systems and signaling cascades. GLP-1 binds the GLP-1 receptor (GLP-1R), a class B Gs-coupled GPCR that increases intracellular cAMP through adenylyl cyclase activation. GLP-1 receptors are expressed on pancreatic beta cells (where they mediate insulin secretion), vagal afferent neurons, area postrema, NTS, and various hypothalamic nuclei including the arcuate nucleus and PVN. PYY(3-36) binds the Y2 receptor, a class A Gi-coupled GPCR that decreases intracellular cAMP and activates GIRK channels. Y2 receptors are concentrated on NPY/AgRP neurons in the arcuate nucleus and on vagal afferent terminals.
Because these receptors engage opposing second messenger systems (cAMP increase versus cAMP decrease) in different cell populations, there is no receptor-level competition or desensitization between GLP-1 and PYY(3-36). A neuron expressing only GLP-1 receptors will respond fully to GLP-1 regardless of PYY levels in its environment, and vice versa. Furthermore, the neurons expressing these receptors are functionally distinct: GLP-1 receptors on POMC neurons directly activate the anorexigenic pathway, while Y2 receptors on NPY/AgRP neurons inhibit the orexigenic pathway. The net result is simultaneous activation of satiety and inhibition of hunger, a "push-pull" mechanism that produces a stronger appetite-suppressing signal than either input alone.
Vagal afferent neurons in the gut wall and nodose ganglion express both GLP-1 receptors and Y2 receptors, creating a potential site of convergence at the peripheral level. When both PYY(3-36) and GLP-1 activate their respective receptors on the same vagal afferent fiber, the resulting signal transmitted to the brainstem is more intense than the signal generated by either hormone alone. This peripheral convergence adds a second level of interaction beyond central nervous system receptor co-activation, further amplifying the combined appetite-suppressing effect.
Preclinical Evidence for GLP-1/PYY Combination Therapy
The translation from physiological observations to therapeutic development has been driven by preclinical studies in rodent models of diet-induced obesity. A 2024 study published in Endocrine Connections investigated the combination of GLP-1 receptor agonists and PYY analogs in diet-induced obese rats. Combined treatment with semaglutide and a non-selective PYY analog produced maximum body weight loss of 14.0 +/- 4.9%, significantly exceeding the 9.9 +/- 1.5% weight loss achieved with semaglutide alone. The combination also produced greater reductions in food intake, fat mass, and hepatic steatosis compared to monotherapy with either agent.
Another preclinical study compared combined PYY(3-36) and liraglutide treatment to Roux-en-Y gastric bypass in diet-induced obese rats. The combination of PYY(3-36) and liraglutide produced weight loss comparable to RYGB surgery and significantly exceeding monotherapy with either compound. This finding is remarkable because it suggests that pharmacological manipulation of the same hormonal axes altered by surgery can replicate surgical weight loss outcomes without operative intervention. The combination also improved glucose tolerance, reduced liver fat, and decreased markers of systemic inflammation, effects that paralleled the metabolic benefits of RYGB.
A triple agonist approach was explored in a study published in Scientific Reports (2023), describing a peptide with agonist activity at GLP-1 receptors, neuropeptide Y1 receptors, and neuropeptide Y2 receptors. This single molecule combined incretin activity with appetite modulation through the NPY receptor system, producing significant weight loss and improved glycemic control in diabetic mouse models. More recently, a rationally designed dual-agonist peptide (bGLP/PYY-19) targeting both GLP-1 and Y2 receptors was reported to significantly outperform semaglutide monotherapy in reducing body weight in diet-induced obese mice. The dual agonist also produced significantly less nausea-associated behavior (measured by pica, a validated rodent model of nausea) compared to semaglutide alone, addressing a critical limitation of current GLP-1 receptor agonist therapy.
Clinical Trials of PYY Analogs with GLP-1 Agonists
Novo Nordisk conducted the most advanced clinical development program combining PYY and GLP-1 receptor agonist therapy. Their long-acting PYY(3-36) analog, NNC0165-1875 (PYY1875), was designed for once-weekly subcutaneous administration as an add-on to semaglutide. PYY1875 incorporated fatty acid acylation to extend its half-life and achieve sustained Y2 receptor activation throughout the dosing interval.
Phase 1 studies in healthy overweight or obese volunteers established the pharmacokinetic profile and dose range of PYY1875 as monotherapy and in combination with semaglutide. All tested dose levels of PYY1875 were tolerated, both alone and when co-administered with semaglutide. The primary adverse events were gastrointestinal in nature, including nausea, decreased appetite (which could be considered both a side effect and a desired therapeutic effect), and abdominal discomfort. Pharmacokinetic analysis confirmed sustained plasma drug levels consistent with once-weekly dosing.
Phase 2 studies evaluated efficacy in participants with overweight or obesity. While PYY1875 demonstrated additional weight loss when combined with semaglutide compared to semaglutide alone, the incremental benefit was modest and was accompanied by a significantly increased rate of gastrointestinal adverse events. The combination produced more nausea, vomiting, and diarrhea than semaglutide alone, reflecting the additive burden on the GI system from engaging both GLP-1 and Y2 receptor-mediated nausea pathways simultaneously.
Novo Nordisk ultimately discontinued development of PYY1875, though the company has not provided detailed rationale beyond citing an unfavorable benefit-risk profile. The decision likely reflects the challenging therapeutic window: the doses of PYY analog needed to produce clinically meaningful additional weight loss on top of semaglutide also produced intolerable GI side effects in a substantial proportion of participants. This mirrors the broader challenge in gut hormone combination therapy, where the mechanisms that suppress appetite (area postrema activation, gastric emptying delay, visceral afferent signaling) also produce the most common and troublesome side effects.
Implications for Current GLP-1 Receptor Agonist Therapy
While dedicated PYY pharmaceutical development has faced setbacks, the understanding of GLP-1/PYY cooperation has practical implications for patients currently using GLP-1 receptor agonists for weight management. The key insight is that endogenous PYY secretion can be optimized through dietary and lifestyle strategies to complement the pharmacological effects of semaglutide, tirzepatide, or liraglutide, potentially enhancing appetite suppression without the side effect burden of exogenous PYY administration.
Dietary strategies that boost PYY work by the same mechanisms outlined earlier: high-protein meals (25-40 grams per meal), adequate fiber intake (25-35 grams daily from diverse sources), and regular meal timing. For patients on GLP-1 receptor agonists who experience reduced appetite, there is a practical tension: the drug reduces hunger and food intake, which can inadvertently lower PYY-stimulating nutrient exposure if meals become too small or protein-poor. Clinical guidance should emphasize that while total caloric intake should decrease on GLP-1 therapy, meal composition should remain protein-rich and fiber-rich to maintain endogenous PYY support. Using the dosing calculator can help patients optimize their overall treatment protocol.
The co-secretion of PYY and GLP-1 from L-cells also means that the same dietary factors that boost PYY also increase endogenous GLP-1 release, creating a double benefit. Patients on exogenous GLP-1 receptor agonists may still benefit from enhanced endogenous GLP-1 because the drug and the native hormone can act at different GLP-1 receptor populations (central versus peripheral) and the endogenous GLP-1 has additional paracrine effects in the gut that the systemically administered drug may not fully replicate. The combined effect of exogenous GLP-1R agonist plus optimized endogenous GLP-1 and PYY release represents an integrated approach that leverages both pharmacological and physiological mechanisms for maximal appetite control.
Future Directions: Multi-Receptor Agonist Design
The discontinuation of PYY1875 has not ended pharmaceutical interest in Y2 receptor-targeting therapeutics. Instead, it has redirected focus toward molecules that combine Y2 agonism with GLP-1 receptor agonism within a single peptide, potentially offering better tolerability through unified pharmacokinetics and reduced peak Y2 receptor activation. Several academic and industry groups are pursuing this approach.
A triple-agonist peptide targeting GLP-1, glucagon, and Y2 receptors has been described, based on the Xenopus GLP-1 peptide scaffold. By incorporating Y2 agonist activity into a molecule that also activates GLP-1 and glucagon receptors, the designers aim to achieve the metabolic benefits of retatrutide-like triple agonism while adding PYY-mediated appetite suppression. Preclinical data in obese mice show greater weight loss with the triple agonist compared to dual GLP-1/glucagon agonists alone, with an acceptable side effect profile.
The dual GLP-1/Y2 agonist peptide bGLP/PYY-19 represents another approach. Designed through rational structural engineering, this peptide combines the GLP-1 receptor-binding domain with a Y2 receptor-binding segment in a single molecule. The critical advantage reported is reduced emesis: by avoiding the high peak Y2 receptor stimulation that occurs with separate PYY analog dosing, the dual agonist may achieve Y2-mediated appetite suppression within the tolerability window. If this finding translates to humans, it would address the primary limitation that sank the PYY1875 program.
These multi-agonist molecules are still in early preclinical development and face significant hurdles before reaching patients, including manufacturing complexity, stability challenges, and the need for extensive safety testing. However, they represent the logical evolution of the gut hormone combination concept and build directly on the PYY-GLP-1 cooperation research. The peptide research hub covers emerging multi-receptor agonist approaches in more detail.
Neural Convergence Points: Where PYY and GLP-1 Meet in the Brain
Understanding the specific brain regions where PYY and GLP-1 signals converge helps explain the additive nature of their combined effects and identifies potential targets for future therapeutic interventions. Functional neuroimaging studies using fMRI in human subjects receiving PYY(3-36) and/or GLP-1 infusions have mapped the neural responses to each hormone individually and in combination, revealing both shared and distinct activation patterns.
In the hypothalamus, PYY(3-36) and GLP-1 activate partially overlapping but distinct neuronal populations. PYY(3-36) primarily affects the arcuate nucleus through Y2 receptor-mediated inhibition of NPY/AgRP neurons, while GLP-1 acts on GLP-1R-expressing neurons in the arcuate nucleus (both NPY/AgRP and POMC populations), paraventricular nucleus (PVN), and dorsomedial hypothalamus (DMH). The PVN is a particularly important convergence point because it receives direct glutamatergic input from arcuate POMC neurons (activated by PYY-mediated NPY/AgRP inhibition) and direct GLP-1R-mediated activation from preproglucagon neurons projecting from the NTS. When both inputs arrive simultaneously, PVN neurons experience stronger net activation, producing a more intense anorexigenic output signal than either input alone.
The brainstem contains another critical convergence zone. The area postrema and adjacent NTS receive both humoral PYY(3-36) (crossing the permeable blood-brain barrier of the area postrema) and GLP-1 (from circulating hormone and from local GLP-1-producing neurons within the NTS itself). Brainstem neurons expressing both Y2 and GLP-1 receptors have been identified, providing a cellular substrate for signal integration. The NTS also receives vagal afferent input carrying both PYY and GLP-1 signals from the gut, adding a third pathway of convergence. This multi-modal brainstem integration explains why combined PYY and GLP-1 exposure produces qualitatively different satiety than either hormone alone: the brainstem generates a more complete and harder-to-override "stop eating" signal when both hormonal channels are simultaneously active.
Cortical convergence occurs in the insular cortex, orbitofrontal cortex (OFC), and anterior cingulate cortex (ACC). These higher-order brain regions process the hedonic and motivational aspects of appetite, determining not just whether you feel full but whether food remains appealing and worth seeking. PYY(3-36) and GLP-1 each independently reduce food-cue-related activation in the OFC and striatum, but combined exposure produces greater suppression than either alone. The insular cortex, which integrates visceral sensory information into conscious awareness of bodily states, shows enhanced activation during combined PYY/GLP-1 exposure, possibly reflecting heightened awareness of satiety signals that promotes conscious recognition of fullness and voluntary meal termination.
Lessons from GLP-1/PYY for Next-Generation Anti-Obesity Therapeutics
The GLP-1/PYY interaction story provides several important lessons for the broader field of anti-obesity drug development. First, it demonstrates that targeting a single pathway, even as effectively as current GLP-1 receptor agonists do, leaves room for improvement through engagement of complementary mechanisms. The 15-22% weight loss achieved by semaglutide and tirzepatide, while impressive, falls short of the 25-35% achievable with bariatric surgery, which engages multiple hormonal pathways simultaneously including both GLP-1 and PYY. This "surgery gap" motivates multi-target therapeutic approaches.
Second, the PYY1875 experience illustrates that combining two separate appetite-suppressing peptides, each with its own side effect profile, can produce intolerable cumulative adverse effects even when each is well-tolerated alone. This argues for molecular integration (single molecules with dual activity) rather than co-administration of separate agents, a lesson being applied in the design of next-generation multi-agonists. The success of tirzepatide (dual GIP/GLP-1 agonism in a single molecule) versus the challenges of separate GIP + GLP-1 co-administration supports this principle.
Third, the additive interaction between PYY and GLP-1 validates the concept of "orthogonal" therapeutic targeting, engaging multiple receptor systems that converge on a common outcome (appetite suppression) through independent mechanisms. This strategy is distinct from "sequential" targeting (hitting the same pathway at multiple points) and offers advantages in terms of reduced risk of tolerance development and cross-resistance. Future anti-obesity therapeutics may combine GLP-1R, Y2R, amylin, and glucagon receptor activities in various combinations, each contributing its unique mechanistic component to a comprehensive appetite and metabolic control strategy.
For current clinical practice, these lessons translate into the recommendation that patients on semaglutide or tirzepatide should actively support their endogenous PYY system through the dietary and lifestyle measures described in this report. While exogenous PYY therapy is not yet available, the "orthogonal" principle means that maximizing endogenous PYY through protein, fiber, exercise, and sleep provides additional appetite control that does not compete with or diminish the effects of the GLP-1 receptor agonist. Instead, it operates through the Y2 receptor pathway to add an independent layer of satiety signaling, approximating (in a much milder form) the multi-hormone effect that makes bariatric surgery so effective.
Bariatric Surgery & PYY

Why Bariatric Surgery Is So Effective: The Hormonal Hypothesis
Bariatric surgery remains the most effective intervention for sustained major weight loss, with Roux-en-Y gastric bypass (RYGB) producing average excess weight loss of 60-75% and sleeve gastrectomy (SG) achieving 50-65% at 5 years. For decades, the efficacy of bariatric surgery was attributed primarily to mechanical restriction (smaller stomach pouch limiting meal size) and, in bypass procedures, malabsorption (bypassed intestinal segment reducing caloric absorption). But these mechanical explanations proved inadequate: RYGB patients consistently achieved greater and more sustained weight loss than restrictive-only procedures with equivalent pouch sizes, and caloric malabsorption after RYGB was found to account for only a small fraction of the caloric deficit needed to explain the observed weight loss.
The hormonal hypothesis, now strongly supported by two decades of research, proposes that bariatric surgery achieves its effects primarily through profound alterations in gut hormone secretion. The anatomical rearrangement of the GI tract in RYGB delivers partially digested nutrients directly from a small gastric pouch to the mid-jejunum, bypassing the duodenum and proximal jejunum. This means nutrients reach the distal small intestine and colon much earlier and in higher concentrations than in normal anatomy, producing dramatically amplified stimulation of the L-cells that populate these regions. The result is massive increases in postprandial PYY and GLP-1 secretion that fundamentally reset appetite regulation.
PYY Changes After Roux-en-Y Gastric Bypass
The changes in PYY secretion after RYGB are among the most dramatic hormonal alterations observed after any surgical procedure. Before surgery, morbidly obese patients typically show low fasting PYY levels and a flat postprandial response, consistent with the PYY deficiency pattern described earlier. Within one week of RYGB, this pattern reverses completely. The area under the curve (AUC) for postprandial PYY increases from approximately 9.9 pg/mL x min to 224.2 pg/mL x min, a greater than 20-fold increase (P = 0.0004). This exaggerated PYY response is detectable within days of surgery, before significant weight loss has occurred, confirming that it is the anatomical rearrangement, not the weight loss itself, that drives the change.
The postprandial PYY peak after RYGB is not only larger but also earlier than before surgery. Pre-operatively, the already blunted PYY response typically peaked at 60-90 minutes after a meal. Post-operatively, peak PYY levels occur at 30-45 minutes, reflecting the rapid delivery of nutrients to L-cell-rich distal intestinal segments through the Roux limb. The peak concentration can reach 3-5 times normal postprandial levels, creating a supraphysiological satiety signal that powerfully suppresses appetite.
Fasting PYY levels show a more complex pattern. Some studies report decreased fasting PYY after RYGB, while others show minimal change. The key finding is the dramatic amplification of the postprandial response, which is the physiologically relevant satiety signal. The ratio of postprandial peak to fasting baseline increases dramatically, meaning the "on-off" signal becomes much more pronounced. This enhanced contrast between low fasting PYY (promoting appropriate meal initiation) and high postprandial PYY (promoting satiety and meal termination) may contribute to the improved eating behavior patterns observed after RYGB.
Longitudinal studies show that the exaggerated PYY response persists long-term, with measurements at 1, 2, 5, and even 10 years after RYGB showing sustained elevation of postprandial PYY. While the very high levels seen in the first few months post-operatively may moderate somewhat over time, the PYY response remains dramatically higher than pre-operative values and substantially higher than in weight-matched never-obese controls. This sustained hormonal change parallels the sustained weight loss that distinguishes bariatric surgery from most dietary and pharmacological weight loss interventions, where weight regain within 1-3 years is the norm.
Cellular Mechanisms: L-Cell Hyperplasia After Surgery
The exaggerated PYY response after RYGB is not solely due to altered nutrient delivery kinetics. Histological studies of intestinal biopsies taken before and after surgery reveal structural changes in the gut mucosa that increase hormone-producing capacity. The Roux limb and common channel (the intestinal segment below the anastomosis where pancreatic and biliary secretions mix with nutrients) undergo mucosal hypertrophy, with increased villus height, crypt depth, and overall mucosal surface area.
Consistent with this general hypertrophy, there is a clear increase in the absolute number of enteroendocrine cells, specifically L-cells. The increase in L-cell density in the Roux limb has been documented in both human studies and animal models of RYGB. This L-cell hyperplasia means that the intestinal segment receiving nutrients after bypass contains more hormone-producing cells per unit length than the same segment before surgery. Combined with the earlier and more concentrated nutrient exposure, the result is a multiplicative effect on total PYY (and GLP-1) secretion.
The stimulus for L-cell hyperplasia appears to involve nutrient sensing and growth factor signaling. GLP-2, co-secreted with GLP-1 and PYY from L-cells, is a potent intestinal growth factor that promotes epithelial proliferation and mucosal expansion. The elevated GLP-2 levels after RYGB likely contribute to the mucosal hypertrophy and L-cell expansion, creating a positive feedback loop: enhanced L-cell stimulation produces more GLP-2, which promotes more L-cell growth, further amplifying hormone output. Changes in bile acid signaling after RYGB, where bile acids enter the alimentary limb earlier and in higher concentrations, may also stimulate L-cell proliferation through TGR5 and FXR receptor activation.
PYY Changes After Sleeve Gastrectomy
Sleeve gastrectomy (SG), which involves removal of approximately 80% of the stomach along the greater curvature without intestinal rearrangement, also produces significant changes in PYY levels, though the pattern differs from RYGB. After SG, postprandial PYY levels increase significantly compared to pre-operative values, but the magnitude of increase is generally less than after RYGB. A meta-analysis comparing the two procedures found that the AUC for postprandial PYY was greater after RYGB than after SG, consistent with the greater weight loss typically achieved with bypass.
The mechanism of increased PYY after SG differs from RYGB because the intestinal anatomy is not altered. The primary explanation is accelerated gastric emptying. The narrow, tubularized stomach remnant after SG empties rapidly, delivering nutrients to the small intestine and subsequently to the distal gut faster than the intact stomach. This faster transit increases nutrient contact with L-cells, stimulating greater PYY and GLP-1 release. Additionally, removal of the gastric fundus eliminates the majority of ghrelin-producing X/A-like cells, dramatically reducing circulating ghrelin. The combination of increased PYY and decreased ghrelin shifts the appetite hormone balance strongly toward satiety.
Comparative studies between RYGB and SG show that at 1 year post-operatively, RYGB patients have approximately 30% greater postprandial PYY levels than SG patients. However, both procedures produce significantly elevated PYY compared to pre-operative baselines, and both produce clinically meaningful weight loss. The difference in PYY elevation between procedures correlates with the difference in weight loss: RYGB typically achieves 3-5% greater total weight loss than SG at long-term follow-up. Whether this weight loss differential is primarily driven by the PYY difference or by other hormonal and anatomical differences between the procedures remains debated.
The Role of PYY in Post-Surgical Appetite Suppression
To establish a causal role for PYY in the appetite suppression observed after bariatric surgery, rather than merely a correlational association, researchers have used pharmacological blockade experiments. In these studies, patients who have undergone RYGB are given specific receptor antagonists to block PYY and/or GLP-1 signaling, and the effect on appetite and food intake is measured.
Blockade of Y2 receptors alone partially reverses the appetite suppression seen after RYGB, increasing food intake compared to the post-surgical baseline. Blockade of GLP-1 receptors alone produces a similar partial reversal. Combined blockade of both Y2 and GLP-1 receptors produces greater appetite restoration than blockade of either receptor alone, though it does not fully restore pre-operative appetite levels. This indicates that PYY and GLP-1 are necessary but not sufficient to explain the complete appetite suppression after RYGB, with other factors including altered bile acid signaling, gut microbiome changes, vagal nerve remodeling, and possible contributions from other gut peptides (oxyntomodulin, glicentin) also playing roles.
A 2025 study published in Obesity Surgery examined the interaction between postprandial exercise and gut hormone release after RYGB. The study found that exercise performed after eating further enhanced the PYY and GLP-1 response in post-RYGB patients, and this enhanced hormonal response was associated with improved insulin sensitivity. This suggests that the combination of surgical anatomy alteration, post-meal PYY/GLP-1 surge, and exercise-induced hormonal amplification creates a particularly favorable metabolic environment in physically active post-surgical patients.
PYY Levels After Failed Bariatric Surgery
Not all bariatric surgery patients achieve or maintain adequate weight loss. Approximately 10-20% of RYGB patients experience insufficient initial weight loss or significant weight regain after an initial successful period. Studies of these "failed" surgical patients reveal that their postprandial PYY levels are lower than those of successful surgical patients, though still higher than pre-operative values. This finding supports the hypothesis that PYY plays a causal role in surgical weight loss outcomes and suggests that inadequate PYY response may be a biomarker for, or contributor to, surgical failure.
A 2025 study examining meal-induced PYY and GLP-1 responses after sleeve gastrectomy found that the magnitude of early postprandial PYY and GLP-1 secretion predicted long-term weight loss outcomes. Patients with the highest PYY and GLP-1 responses at 3 months post-surgery achieved the greatest weight loss at 1, 2, and 3 years. This raises the possibility that postprandial gut hormone testing could identify patients at risk for surgical failure early in the post-operative course, allowing targeted interventions such as dietary optimization, exercise programs, or adjunctive pharmacotherapy with GLP-1 receptor agonists to support these patients.
For patients who experience weight regain after bariatric surgery, addition of semaglutide or tirzepatide has shown promise in clinical practice. The rationale is straightforward: these medications provide exogenous GLP-1 receptor activation to supplement the enhanced but potentially insufficient endogenous GLP-1 and PYY responses after surgery. The combination of surgical hormonal changes plus pharmacological GLP-1R agonism can produce total appetite suppression greater than either approach alone, helping patients return to a weight loss trajectory.
Comparing Bariatric Surgery PYY Effects with Pharmacological Approaches
The dramatic PYY elevations after bariatric surgery set a high bar for pharmacological replication. Current GLP-1 receptor agonists do not directly increase PYY levels; they provide exogenous activation of GLP-1 receptors while PYY signaling depends entirely on endogenous L-cell secretion. The most potent available weight loss medications, semaglutide 2.4 mg weekly and tirzepatide at maximum dose, produce average weight loss of 15-22%, approaching but not matching the 25-35% weight loss achievable with RYGB.
This "efficacy gap" between surgery and drugs has motivated the development of combination approaches that more closely replicate the full hormonal profile of post-surgical physiology. The theoretical ideal would combine GLP-1R agonism (replicating the surgical GLP-1 increase) with Y2 receptor agonism (replicating the surgical PYY increase) and possibly glucagon receptor agonism (replicating the enhanced energy expenditure and hepatic fat reduction seen after surgery). Retatrutide, a triple GLP-1/GIP/glucagon receptor agonist, has achieved average weight loss of 24% in clinical trials, approaching surgical levels. Adding Y2 receptor agonism to such a molecule could potentially close the remaining gap, though the tolerability challenges described earlier must be overcome.
The pharmacological replication of post-surgical gut hormone profiles represents one of the most active areas of anti-obesity drug development. While no current commercial product includes a PYY or Y2 receptor agonist component, the preclinical pipeline suggests that future combination therapies may incorporate this axis. The drug comparison hub provides updated comparisons of available weight management medications, including their effects on gut hormone profiles.
Bile Acid Changes and Their Impact on PYY After Surgery
Bariatric surgery produces significant alterations in bile acid metabolism that contribute to enhanced PYY secretion through mechanisms independent of nutrient delivery changes. In normal anatomy, bile is secreted from the gallbladder into the duodenum, where it mixes with food and facilitates fat digestion and absorption. After RYGB, bile enters the biliopancreatic limb (the bypassed duodenum) and does not contact food until the jejunojejunal anastomosis, where the alimentary and biliopancreatic limbs join. This altered bile flow means that bile acids reach the distal gut in higher concentrations and altered molecular composition compared to pre-operative physiology.
Bile acids are potent activators of the TGR5 (Takeda G protein-coupled receptor 5, also known as GPBAR1) on L-cells. TGR5 activation increases intracellular cAMP and triggers PYY and GLP-1 secretion. After RYGB, total bile acid levels increase significantly, and the bile acid pool shifts toward more hydrophilic species with greater TGR5 agonist activity. Studies measuring bile acid profiles before and after RYGB show 2-3 fold increases in total plasma bile acids, with disproportionate increases in the TGR5-active conjugated bile acid species. This bile acid-mediated L-cell stimulation adds to the nutrient-contact and L-cell hyperplasia mechanisms, creating a third pathway through which surgery enhances PYY output.
The bile acid-PYY connection has therapeutic implications beyond surgery. Bile acid sequestrants (colesevelam, cholestyramine) alter the enterohepatic circulation of bile acids, increasing colonic bile acid exposure and potentially stimulating L-cell PYY release. While these drugs are primarily used for cholesterol lowering and glycemic control in type 2 diabetes, their ability to modulate PYY through bile acid-TGR5 signaling represents a pharmacological avenue for PYY augmentation that could complement GLP-1 receptor agonist therapy. Ursodeoxycholic acid (UDCA), a secondary bile acid with TGR5 agonist properties, has been studied as a potential metabolic therapeutic, though results on PYY levels have been mixed.
Long-term Outcomes: PYY's Role in Weight Maintenance After Surgery
One of the most remarkable features of bariatric surgery is the durability of weight loss, with most RYGB patients maintaining 50-60% excess weight loss at 10 years and beyond. This stands in stark contrast to dietary weight loss, where the majority of lost weight is typically regained within 2-5 years. The sustained elevation of PYY and GLP-1 after surgery is believed to be a key mechanism behind this weight loss maintenance, essentially providing permanent hormonal support for reduced appetite and lower caloric intake.
Longitudinal hormonal studies tracking PYY levels from before surgery through 10+ years post-operatively show a consistent pattern. Postprandial PYY peaks dramatically in the first months after RYGB, then moderates somewhat over the first 1-2 years as the gut adapts, but stabilizes at levels 3-5 times higher than pre-operative values. This sustained elevation means that patients continue to experience enhanced meal-induced satiety for years after surgery, providing an ongoing biological mechanism for appetite control that does not depend on willpower, dietary discipline, or continued medical intervention.
The contrast with weight regain is informative. Among the minority of RYGB patients who experience significant weight regain, postprandial PYY levels tend to decline toward pre-operative values, though they rarely return completely to the pre-surgical baseline. Whether the PYY decline causes the weight regain or reflects other changes (dilated gastrojejunal anastomosis, dietary pattern deterioration, psychological factors) is debated. However, the association between declining PYY and weight regain supports PYY as a key mediator of surgical success and identifies PYY monitoring as a potential tool for early detection of patients at risk for weight recidivism.
For patients who have undergone bariatric surgery and are experiencing weight regain, the addition of semaglutide or tirzepatide provides exogenous GLP-1 receptor stimulation to supplement the declining endogenous response. Simultaneously, dietary optimization to support endogenous PYY (high protein, high fiber, regular meals) can help maintain or restore the PYY-mediated component of appetite control. This combined pharmacological and dietary approach attempts to recreate the hormonal milieu that characterized the initial post-surgical weight loss period, potentially stabilizing weight and preventing further regain.
PYY in Emerging Bariatric Procedures
Newer bariatric procedures and endoscopic interventions have been developed with an increasingly sophisticated understanding of gut hormone physiology, including PYY. Endoscopic sleeve gastroplasty (ESG), which uses suturing to reduce gastric volume without surgical incision, produces modest increases in postprandial PYY compared to the dramatic changes seen with RYGB, consistent with its more moderate weight loss outcomes (15-20% total body weight loss versus 25-35% with RYGB). The degree of PYY elevation after ESG correlates with weight loss success, supporting the hormone's role as a mechanistic mediator rather than merely a biomarker.
Duodenal mucosal resurfacing (DMR), a catheter-based procedure that ablates the duodenal mucosa to alter nutrient sensing and hormone signaling, has been investigated for its effects on gut hormones. Early data suggest that DMR may increase postprandial PYY and GLP-1, though the magnitude is smaller than with RYGB. The one-anastomosis gastric bypass (OAGB, also called mini gastric bypass) produces PYY elevations comparable to RYGB, consistent with similar weight loss outcomes. Biliopancreatic diversion with duodenal switch (BPD-DS), the most malabsorptive bariatric procedure, produces the highest postprandial PYY levels of any surgery, which may contribute to its position as the most effective procedure for long-term weight loss in patients with super-obesity (BMI > 50).
These procedural comparisons reinforce the relationship between intestinal anatomy, PYY secretion, and weight loss outcomes. Procedures that deliver nutrients more rapidly to distal L-cells, increase L-cell mass through intestinal adaptation, or enhance bile acid-mediated L-cell stimulation produce proportionally greater PYY responses and proportionally more weight loss. This consistent relationship across diverse surgical and endoscopic approaches provides strong evidence that PYY is not merely associated with but mechanistically contributes to the appetite regulation that drives bariatric weight loss.
Therapeutic Development
History of PYY Drug Development
The development of PYY(3-36) as a therapeutic agent began in earnest following the Batterham et al. (2002) publication demonstrating its appetite-suppressing effects in humans. Several pharmaceutical and biotechnology companies initiated PYY drug programs in the early 2000s, attracted by the compound's potent anorexigenic activity and favorable mechanism as a natural gut hormone. However, the path from promising biology to approved therapeutic has proven difficult, and no PYY-based drug has yet reached the market.
Nastech Pharmaceutical (later Mdrna) developed an intranasal PYY(3-36) formulation that advanced to Phase 2 clinical trials in the mid-2000s. The intranasal route offered potential advantages including rapid absorption, avoidance of first-pass hepatic metabolism, and patient convenience. Phase 1 studies showed that intranasal PYY(3-36) achieved measurable plasma levels and produced dose-dependent appetite suppression. However, Phase 2 results were disappointing: the intranasal formulation produced inconsistent bioavailability between subjects and administrations, and the appetite-suppressing effects did not reliably translate into significant weight loss over the 12-week study period. The program was discontinued.
7TM Pharma (later Helsinn) pursued Y2 receptor-selective small molecule agonists as an alternative to peptide-based PYY analogs. Small molecules offer potential advantages in oral bioavailability, manufacturing cost, and formulation flexibility. However, the Y2 receptor proved difficult to target with small molecules due to the large, shallow peptide-binding pocket typical of class A GPCRs in the NPY receptor family. Despite extensive medicinal chemistry efforts, no small molecule Y2 agonist achieved sufficient potency and selectivity to advance to clinical development.
Novo Nordisk's PYY Analog Programs
Novo Nordisk, using its expertise in peptide drug design from the development of semaglutide and liraglutide, pursued the most advanced PYY analog programs. Two compounds advanced to clinical testing: NN9748 and NNC0165-1875 (PYY1875).
NN9748 was an early-generation long-acting PYY(3-36) analog that completed Phase 1 studies. The compound incorporated amino acid substitutions and a fatty acid side chain to extend plasma half-life and permit less frequent dosing. Phase 1 data demonstrated adequate pharmacokinetics and a dose-dependent effect on appetite markers, but the program appears to have been superseded by the more advanced PYY1875 molecule.
PYY1875 (NNC0165-1875) represented Novo Nordisk's lead PYY therapeutic candidate, designed specifically as a once-weekly add-on to semaglutide for chronic weight management. The molecule was engineered with C18 fatty acid acylation to promote albumin binding, extending the elimination half-life sufficiently for weekly dosing. Amino acid modifications at key cleavage sites improved resistance to aminopeptidases and carboxypeptidases that degrade native PYY(3-36) in vivo.
The clinical development of PYY1875 progressed through a logical sequence. In diet-induced obese rats, combination of PYY1875 with semaglutide produced weight loss of approximately 14%, significantly exceeding semaglutide monotherapy (approximately 10%). The combination also produced greater fat mass reduction and improvement in liver fat content. These preclinical results supported advancement to human studies.
Phase 1 studies in healthy overweight and obese volunteers demonstrated that PYY1875 was tolerable at all tested dose levels, both as monotherapy and in combination with semaglutide. Pharmacokinetic analysis confirmed sustained plasma concentrations consistent with the target weekly dosing frequency. Exploratory efficacy endpoints showed trends toward greater appetite suppression and food intake reduction with the combination compared to semaglutide alone.
Phase 2 studies evaluated multiple doses of PYY1875 co-administered with semaglutide over longer treatment periods in participants with overweight or obesity. While the combination produced additional weight loss beyond semaglutide alone, the increment was modest and came at the cost of significantly increased gastrointestinal adverse events, particularly nausea, vomiting, and diarrhea. The gastrointestinal side effect profile was more severe with the combination than with semaglutide alone, and the discontinuation rate due to adverse events was higher in the combination groups.
Novo Nordisk announced discontinuation of PYY1875 development, citing an overall benefit-risk assessment that did not support further advancement. The company's public communications indicated that the additional weight loss provided by PYY1875 was insufficient to justify the incremental side effect burden. This decision was made in the context of Novo Nordisk's broader portfolio, which includes cagrilintide (a long-acting amylin analog) as an alternative combination partner for semaglutide, with CagriSema demonstrating approximately 25% weight loss in clinical trials with an acceptable tolerability profile.
Alternative Approaches to Y2 Receptor Targeting
The challenges encountered with dedicated PYY analogs have prompted exploration of alternative strategies for engaging Y2 receptor signaling in obesity treatment. These approaches aim to capture the appetite-suppressing benefits of Y2 activation while avoiding the tolerability limitations of high-dose exogenous PYY analog administration.
Multi-receptor agonist peptides incorporating Y2 activity alongside GLP-1 and/or glucagon receptor agonism represent the most actively pursued alternative. By combining Y2 agonism with GLP-1 receptor agonism in a single molecule, these hybrid peptides may achieve lower peak Y2 receptor activation compared to a separate PYY analog, potentially staying below the nausea threshold while still providing meaningful appetite suppression. The pharmacokinetic advantages are also relevant: a single molecule ensures coordinated exposure of both targets, avoiding the variable PK profiles that can occur when two separate peptide drugs are co-administered.
Gene therapy approaches to increase endogenous PYY production have been explored in preclinical settings. Adeno-associated virus (AAV) vectors encoding PYY have been delivered to the hypothalamus or salivary glands in rodent models, producing sustained PYY overexpression and reduced food intake over weeks to months. While gene therapy for obesity remains far from clinical reality, these studies provide proof of concept that chronic moderate elevation of PYY, as opposed to the acute high-level exposure from exogenous analog administration, can produce weight loss without significant aversion or nausea.
Modulation of endogenous PYY secretion through gut-targeted mechanisms offers another strategy. Encapsulated nutrients designed to survive gastric and proximal small intestinal digestion and release their contents in the distal ileum and colon could directly stimulate L-cells, increasing endogenous PYY (and GLP-1) release without the need for exogenous peptide administration. Several nutraceutical companies have pursued this concept using enteric-coated capsules containing lauric acid, palmitoylethanolamide, or other L-cell secretagogues. Early clinical data show modest increases in postprandial PYY and GLP-1 and small but statistically significant effects on appetite, though the magnitude of effect is much smaller than pharmacological PYY analogs or GLP-1 receptor agonists.
PYY and the Gut Microbiome: An Emerging Therapeutic Interface
The gut microbiome's role in PYY regulation represents a relatively new but rapidly growing area of research with potential therapeutic implications. Certain bacterial species in the colon produce short-chain fatty acids (SCFAs) that directly stimulate L-cell PYY secretion through FFAR2 and FFAR3 receptors. The composition and metabolic activity of the gut microbiome therefore influence PYY levels and, by extension, appetite regulation.
Obesity is associated with altered gut microbiome composition (dysbiosis), characterized by reduced microbial diversity and changes in the relative abundance of specific phyla and species. Some of these changes reduce SCFA production, potentially contributing to the reduced PYY levels seen in obesity. Conversely, interventions that restore microbial diversity and SCFA production, including prebiotic fiber supplementation, probiotic administration, and fecal microbiota transplantation, have been shown to modestly increase PYY levels in both animal models and human studies.
Bariatric surgery produces dramatic changes in gut microbiome composition, with increased abundance of Gammaproteobacteria, Verrucomicrobia (particularly Akkermansia muciniphila), and Fusobacterium, and decreased Firmicutes-to-Bacteroidetes ratio. These microbial changes may contribute to the enhanced PYY secretion after surgery through increased SCFA production and altered bile acid metabolism. The concept of microbiome modulation as a strategy to enhance PYY levels is particularly attractive because it could complement pharmacological approaches without adding to the gastrointestinal side effect burden that limits PYY analog therapy.
PYY in Type 2 Diabetes Therapeutics
Beyond its role in appetite and body weight regulation, PYY has attracted attention as a potential therapeutic target for type 2 diabetes. PYY influences glucose homeostasis through multiple mechanisms: it slows gastric emptying (reducing the rate of postprandial glucose absorption), modulates insulin secretion, and enhances peripheral insulin sensitivity. A study demonstrating that subcutaneous infusion of three hormones (GLP-1, PYY, and oxyntomodulin) over a 4-week period improved glucose tolerance in diabetic patients to a degree comparable to or exceeding RYGB surgery represents a compelling proof of concept for multi-hormone metabolic therapy.
However, unlike GLP-1, PYY does not have a direct insulinotropic effect. It does not stimulate insulin secretion from beta cells in a glucose-dependent manner. Its glycemic benefits are indirect, operating through appetite and weight reduction, gastric emptying modulation, and peripheral insulin sensitization. This means PYY is unlikely to serve as a standalone diabetes therapy but could provide additive metabolic benefits when combined with agents that directly enhance insulin secretion, such as semaglutide or tirzepatide.
The observation that PYY deficiency precedes the development of type 2 diabetes in genetically susceptible individuals, appearing even before insulin resistance and adiposity, raises the possibility that early PYY augmentation could prevent or delay diabetes onset. This preventive application has not been tested in clinical trials but represents a conceptually distinct indication from the treatment of established obesity or diabetes.
Challenges and Opportunities in PYY Therapeutics
Several cross-cutting challenges have slowed PYY therapeutic development and will need to be addressed for future success. First, the narrow therapeutic window between effective appetite suppression and intolerable nausea remains the central limitation. Any successful PYY-targeting therapy must achieve sufficient Y2 receptor activation to meaningfully reduce food intake without crossing the nausea threshold, a challenge compounded when the drug is combined with GLP-1 receptor agonists that have their own nausea liability.
Second, the short half-life of native PYY(3-36) necessitates complex chemical modifications for long-acting formulations, which can alter receptor binding kinetics, tissue distribution, and immunogenicity. Balancing pharmacokinetic extension with preserved pharmacological activity is an ongoing engineering challenge. Third, the lack of selective Y2 agonist small molecules limits the therapeutic modalities available. Peptide drugs require parenteral administration, which is less convenient than oral dosing and limits the addressable patient population.
Opportunities exist in several areas. The strong preclinical rationale for GLP-1/Y2 combination therapy remains valid despite the clinical failure of PYY1875 as a separate add-on. Single-molecule dual agonists may overcome the tolerability issues by providing coordinated, moderate activation of both targets. Gut-directed strategies to enhance endogenous PYY, including microbiome modulation and ileal-targeted nutrient delivery, offer low-risk, potentially complementary approaches to complement existing pharmacotherapies. And the growing understanding of PYY's role in hedonic appetite regulation opens possibilities for targeting specific eating behaviors (binge eating, emotional eating) where current therapies have limited efficacy.
For patients and clinicians today, the practical takeaway is that PYY biology can be leveraged through dietary and lifestyle strategies even without dedicated PYY drugs. High-protein, high-fiber diets, regular aerobic exercise, adequate sleep, and stress management all support endogenous PYY secretion and complement the effects of available GLP-1-based weight management therapies. The lifestyle hub provides detailed guidance on integrating these approaches into a comprehensive weight management plan.
Current Clinical Landscape
No PYY-based therapeutic is currently approved or in late-stage clinical development. However, PYY biology is central to understanding why bariatric surgery works, why GLP-1 receptor agonists are effective, and how future multi-hormone therapies may close the efficacy gap between medications and surgery. Patients using semaglutide, tirzepatide, or liraglutide can optimize endogenous PYY through dietary protein and fiber intake, regular exercise, and consistent meal timing.
Practical PYY Optimization & Dietary Strategies
While no PYY-based drug is currently available, the biology of PYY secretion is well understood enough that individuals can meaningfully boost their endogenous PYY levels through targeted dietary, exercise, and lifestyle strategies. For people already using GLP-1 receptor agonists for weight management, optimizing PYY output may provide additional appetite control that complements the drug's effects, potentially allowing lower doses or improved weight loss results.
Dietary Triggers for PYY Release
PYY is released by L-cells in the distal intestine in response to nutrient contact. Not all nutrients trigger PYY release equally, and understanding which dietary components are the strongest PYY stimulants allows for strategic meal design that maximizes natural satiety signaling.
Protein is the strongest PYY trigger. Protein-rich meals produce 30-50% higher PYY responses than isocaloric carbohydrate or fat meals. The mechanism involves both direct amino acid sensing by L-cells and indirect effects through gastric peptides stimulated by protein digestion. Whey protein appears to be particularly potent for PYY stimulation, possibly due to its rapid digestion kinetics and specific amino acid profile. Practical application: frontloading protein at the beginning of meals and ensuring 25-40 grams of protein per meal optimizes PYY output throughout the day.
Dietary fiber, particularly soluble fiber, stimulates PYY release through two mechanisms. First, fiber slows gastric emptying, prolonging nutrient contact time in the ileum where L-cells are most concentrated. Second, fermentation of fiber by colonic bacteria produces short-chain fatty acids (SCFAs) that directly stimulate L-cell PYY secretion through free fatty acid receptors (FFAR2 and FFAR3). This second mechanism explains why regular fiber intake improves PYY responses over time: it takes days to weeks for the microbiome to upregulate SCFA production in response to increased fiber intake. Sources with the strongest evidence for PYY stimulation include oat beta-glucan, psyllium, inulin, resistant starch (found in cooled cooked potatoes and rice), and legumes.
Medium-chain triglycerides (MCTs) and certain fatty acids also stimulate PYY. MCTs are absorbed more rapidly than long-chain fatty acids and reach the ileum more quickly, producing earlier PYY responses. Oleic acid (found abundantly in olive oil and avocados) has demonstrated PYY-stimulating effects in human studies. Including healthy fats in meals, particularly those high in oleic acid and MCTs, supports PYY secretion while providing satiation through other mechanisms as well.
Meal timing and frequency affect PYY patterns. PYY exhibits a cumulative pattern across the day, with each meal building on the PYY elevation from the previous one. This means that breakfast produces the lowest absolute PYY response, while dinner (after a full day of eating) produces the highest. Skipping breakfast doesn't just eliminate the morning PYY boost; it reduces the cumulative PYY trajectory for the entire day. For individuals using semaglutide or tirzepatide who find their appetite reduced to the point of skipping meals, this is an important consideration: eating regular meals, even if smaller, maintains PYY cycling patterns that support sustained appetite control between doses.
Exercise and PYY
Aerobic exercise acutely stimulates PYY release, an effect that helps explain the temporary appetite suppression many people experience after vigorous workouts (sometimes called "exercise anorexia"). The intensity threshold appears to be around 60-70% of maximum heart rate, with more vigorous exercise producing stronger PYY responses. Duration also matters: exercise sessions of 30 minutes or longer produce more sustained PYY elevation than shorter bouts.
High-intensity interval training (HIIT) produces particularly strong PYY responses, exceeding those from moderate continuous exercise matched for total caloric expenditure. The intermittent surges in blood flow to the intestinal vasculature during HIIT may stimulate L-cells more effectively than the sustained but lower-intensity stimulation of steady-state exercise. For individuals using GLP-1 therapy who want to maximize natural appetite-suppressing mechanisms between doses, scheduling HIIT sessions on days when GLP-1 effects are waning (typically 5-7 days after weekly injection) provides complementary coverage.
Resistance training's effects on PYY are less studied but appear to be more modest than aerobic exercise. However, resistance training's long-term effects on body composition (increased lean mass, decreased fat mass) improve baseline PYY responsiveness over time. Obese individuals consistently show blunted PYY responses compared to lean individuals, and as body composition improves through any means, PYY signaling tends to normalize. This creates a positive feedback loop: better body composition leads to better PYY responses, which leads to better appetite control, which supports continued body composition improvement.
Microbiome Optimization for PYY
The gut microbiome influences PYY secretion primarily through SCFA production. Butyrate, propionate, and acetate, the three main SCFAs produced by bacterial fermentation of dietary fiber, all stimulate L-cell PYY release. The composition of the microbiome determines how efficiently dietary fiber is converted to SCFAs, which means that two people eating identical high-fiber diets may get very different PYY responses based on their microbial populations.
Strategies to optimize the microbiome for PYY production include: gradually increasing fiber intake to 30-50 grams per day (sudden large increases cause bloating and gas as the microbiome adjusts), consuming a diverse range of fiber types (different bacteria ferment different substrates), including fermented foods (yogurt, kefir, sauerkraut, kimchi) to introduce beneficial organisms, and considering a targeted probiotic containing Akkermansia muciniphila, a bacterium specifically associated with improved metabolic signaling and gut hormone production.
For individuals on GLP-1 receptor agonists, microbiome optimization serves a dual purpose: it enhances endogenous PYY production and may reduce some GI side effects. The slower gastric emptying caused by GLP-1 drugs can disrupt normal gut motility patterns, and a strong, diverse microbiome appears to buffer against the dysbiosis that can result from altered intestinal transit. Visit the Lifestyle Hub for comprehensive guidance on nutrition strategies that support gut health and metabolic function.
Sleep, Stress, and PYY
Sleep deprivation significantly reduces postprandial PYY responses. Studies comparing PYY levels after normal sleep (7-8 hours) versus restricted sleep (4-5 hours) show 20-30% lower PYY responses in the sleep-deprived condition. This blunted satiety signaling contributes to the well-documented increase in caloric intake that accompanies sleep loss. For anyone pursuing weight management, whether through lifestyle changes alone or in combination with semaglutide, tirzepatide, or other pharmacotherapy, sleep optimization is not optional. It directly affects the hormonal environment that determines appetite control. Pinealon and DSIP are peptide options for individuals who need sleep architecture support alongside their weight management protocol.
Chronic stress similarly impairs PYY secretion while elevating cortisol-driven appetite signals. The combination of reduced PYY (less satiety) and elevated cortisol (more hunger, particularly for calorie-dense comfort foods) creates a metabolic environment that strongly favors weight gain. Stress management practices, including regular exercise, meditation, social connection, and when appropriate, adaptogenic support with peptides like Selank, help maintain the neuroendocrine environment that allows PYY to function effectively.
Multi-Hormone Approaches & Future Therapeutic Landscape
The most effective weight management interventions, both surgical and pharmacological, work by engaging multiple gut hormone pathways simultaneously rather than relying on a single mechanism. Understanding PYY's role within this multi-hormone framework is essential for appreciating both current therapeutic options and where the field is heading.
The Gut Hormone Orchestra: Why Multiple Signals Matter
Appetite regulation involves a complex interplay of at least six major gut hormones: GLP-1, PYY, GIP, CCK, oxyntomodulin, and ghrelin. Each provides distinct information about nutritional status, and the brain integrates these signals to determine hunger, satiety, food reward, and metabolic rate. When only one of these signals is pharmacologically enhanced (as with pure GLP-1 receptor agonists), the brain receives a conflicting mix of one strong satiety signal and five unchanged or compensatory signals. This may partly explain why weight loss plateaus with monotherapy and why some patients develop tolerance over time.
Bariatric surgery, which remains the gold standard for sustained weight loss, simultaneously alters the secretion of multiple hormones: GLP-1 and PYY increase dramatically (often 5-10 fold), ghrelin decreases, and the timing and magnitude of GIP, CCK, and oxyntomodulin responses all change. The pharmacological challenge is replicating this multi-hormone symphony with drugs. The evolution from single-agonist drugs (liraglutide) to dual-agonists (tirzepatide, targeting GLP-1 and GIP) to triple-agonists (retatrutide, targeting GLP-1, GIP, and glucagon) represents steps toward this multi-hormone ideal.
PYY is the notable absent partner in currently available pharmacotherapy. Adding Y2 receptor agonism to existing GLP-1-based treatments could address the remaining gap between surgical and pharmacological outcomes. The fact that the PYY1875 clinical trial failed doesn't mean the concept is wrong; it means the specific execution (adding a separate PYY infusion to an existing GLP-1 drug) wasn't the right approach. Single-molecule polypharmacology, where one drug activates both GLP-1 and Y2 receptors, may succeed where combination therapy didn't.
Amylin Analogs: The Current Frontier
While PYY-specific drugs remain in development, amylin analogs have emerged as the next major addition to the weight management toolkit. Amylin, co-secreted with insulin from pancreatic beta cells, works through distinct appetite-suppressing pathways in the area postrema and brainstem that complement GLP-1's hypothalamic effects. Cagrilintide, a long-acting amylin analog developed by Novo Nordisk, has shown promising weight loss results alone and in combination with semaglutide.
The combination of cagrilintide plus semaglutide (branded CagriSema) is currently in Phase 3 clinical trials and represents one of the most anticipated weight loss drugs in development. Early data suggest weight loss approaching surgical outcomes, roughly 22-25% total body weight loss in some studies. While this combination doesn't include a PYY component, the cagrilintide-induced slowing of gastric emptying and delayed intestinal transit may indirectly enhance endogenous PYY secretion by prolonging nutrient contact time with ileal L-cells.
For individuals currently using GLP-1 monotherapy who want additional appetite suppression, optimizing endogenous PYY production through the dietary and lifestyle strategies discussed above provides the closest available approximation of PYY pharmacotherapy. The combination of a GLP-1 agonist providing one pathway of satiety signaling, plus dietary optimization maximizing endogenous PYY as a second pathway, plus exercise further boosting PYY while improving metabolic health, creates a multi-mechanism approach using currently available tools.
Peptide Approaches to Metabolic Support
Beyond gut hormones and appetite signaling, several peptides target metabolic pathways that influence weight management through different mechanisms. These don't directly involve PYY but address related aspects of the metabolic equation.
Tesofensine is a triple monoamine reuptake inhibitor originally developed for neurodegenerative diseases that was found to produce significant weight loss in clinical trials. Its mechanism, enhancing dopamine, norepinephrine, and serotonin signaling in appetite-regulating brain circuits, is distinct from and potentially complementary to gut hormone-based approaches. While PYY reduces appetite through peripheral-to-central signaling, tesofensine modulates the central processing of those signals. The combination of enhanced gut-derived satiety signals (from PYY optimization and GLP-1 therapy) with improved central processing of those signals (from tesofensine) could theoretically produce stronger appetite suppression than either approach alone.
5-Amino-1MQ takes yet another approach, inhibiting the enzyme NNMT (nicotinamide N-methyltransferase) to increase cellular energy expenditure. Rather than reducing appetite, it increases the metabolic rate at which consumed calories are burned. This "energy expenditure" approach addresses the other side of the energy balance equation from PYY's appetite-suppressing role. For comprehensive weight management, addressing both sides, eating less (GLP-1, PYY optimization, appetite modulation) and burning more (exercise, 5-Amino-1MQ, metabolic optimization), produces better outcomes than either approach alone.
AOD-9604 and Fragment 176-191 are growth hormone fragments that stimulate lipolysis (fat breakdown) without the growth-promoting effects of full-length GH. These compounds may help mobilize fat stores that PYY and GLP-1 agonists help create through caloric deficit, addressing the mechanical process of fat utilization that must accompany reduced intake for actual body composition change.
The MOTS-c peptide influences metabolism at the mitochondrial level, improving glucose handling and fat oxidation. In animal studies, MOTS-c prevents diet-induced obesity and improves insulin sensitivity. Its metabolic effects complement PYY's appetite effects: MOTS-c helps the body process nutrients more efficiently while PYY helps regulate how much nutrient intake occurs in the first place.
The Integrated Weight Management Framework
Viewing PYY within the broader weight management landscape reveals that the most effective approaches layer multiple interventions across different mechanisms rather than relying on any single agent. A comprehensive framework might include:
Pharmacological foundation: GLP-1 receptor agonist (semaglutide or tirzepatide) for primary appetite suppression and metabolic improvement. Use the FormBlends dosing calculator for personalized titration guidance.
Endogenous hormone optimization: Dietary strategies to maximize PYY, CCK, and amylin secretion. High-protein, high-fiber meals with healthy fats. Consistent meal timing to support cumulative PYY patterns.
Metabolic support: Exercise program combining HIIT (for acute PYY stimulation and cardiovascular fitness) with resistance training (for lean mass maintenance during weight loss). Sleep optimization to maintain normal PYY responsiveness and hormonal balance.
Targeted peptide supplementation: Based on individual needs, specific metabolic bottlenecks, and health goals. The FormBlends free assessment helps identify which compounds are most relevant for individual circumstances.
PYY may not yet have its own drug on pharmacy shelves, but its biology informs every aspect of effective weight management. Understanding how PYY fits into the hormonal orchestra of appetite regulation helps individuals and clinicians make better decisions about diet, exercise, medication timing, and complementary strategies. As multi-hormone therapies advance through clinical development, PYY is likely to find its pharmacological moment, not as a standalone treatment but as one voice in the increasingly sophisticated symphony of metabolic medicines. Visit the GLP-1 Research Hub for comprehensive coverage of the evolving weight management therapeutic landscape.
Special Populations & Individualized PYY Considerations
PYY responses vary significantly across different populations, and understanding these differences helps tailor weight management strategies to individual circumstances. Age, sex, body composition, genetic background, and health conditions all influence PYY secretion, receptor sensitivity, and the overall contribution of PYY to appetite control.
PYY in Aging Populations
Aging affects PYY biology in complex ways. Some studies suggest that older adults actually have higher baseline PYY levels than younger individuals, which may contribute to the age-related anorexia ("anorexia of aging") that puts elderly individuals at risk for malnutrition and sarcopenia. However, the responsiveness of PYY secretion to meals appears to decrease with age, and the central sensitivity to PYY's satiety signal may also change. This creates an unusual situation where baseline appetite suppression increases (through elevated tonic PYY) while meal-specific satiety signaling weakens.
For older adults pursuing weight management, the goal is usually to reduce visceral fat and improve metabolic health while preserving lean muscle mass, a more nuanced objective than simple weight loss. The PYY-related dietary strategies discussed earlier, particularly high-protein meals, are doubly important for this population: protein stimulates PYY for appetite regulation and provides the amino acids needed to maintain muscle mass during caloric deficit. Aiming for 1.2-1.6 grams of protein per kilogram of body weight distributes adequate PYY stimulation across meals while supporting lean tissue preservation.
Growth hormone optimization with compounds like tesamorelin or sermorelin becomes relevant in older adults pursuing body composition change. GH decline with age (somatopause) impairs lipolysis and muscle protein synthesis, making body composition optimization more difficult regardless of appetite control. Combining GLP-1 therapy (for appetite and metabolic control) with GH optimization (for body composition effects) addresses two distinct age-related metabolic declines simultaneously.
Sex Differences in PYY Response
Women generally show higher baseline PYY levels and stronger postprandial PYY responses than men, even after adjusting for body weight and composition. This may partially explain why women tend to eat smaller meals than men and why some weight management strategies work differently between sexes. Estrogen appears to enhance PYY secretion, and the decline in estrogen during menopause may contribute to the appetite changes and weight gain commonly experienced during this transition.
For women in perimenopause or menopause experiencing increased appetite and weight gain, the declining PYY response is one of several hormonal shifts contributing to the problem. Estrogen's effects on PYY secretion, combined with changes in ghrelin sensitivity, insulin dynamics, and cortisol patterns, create a metabolic environment that strongly favors weight gain. Semaglutide or tirzepatide can provide pharmacological appetite control during this transition, while dietary strategies that maximize endogenous PYY production help maintain natural satiety signaling that hormonal changes are eroding.
During pregnancy, PYY levels decrease during the first trimester (allowing increased food intake to support fetal growth) and gradually rise during the third trimester. These physiological PYY changes should not be pharmacologically overridden. GLP-1 receptor agonists are contraindicated during pregnancy, and no PYY-modifying interventions should be attempted during pregnancy or lactation.
PYY After Bariatric Surgery
Understanding PYY's role after bariatric surgery is clinically relevant because it explains both the surgery's effectiveness and potential long-term challenges. Roux-en-Y gastric bypass produces the most dramatic PYY elevation of any intervention, with postprandial PYY levels increasing 5-10 fold within months of surgery. This massive PYY surge is a primary mechanism behind the appetite suppression and metabolic improvement that make RYGB the most effective weight loss intervention available.
Sleeve gastrectomy also increases PYY, though less dramatically than RYGB. The direct removal of the gastric fundus (which produces ghrelin) combined with faster nutrient delivery to the ileum (stimulating more PYY) creates a favorable appetite hormone profile. However, PYY elevation after sleeve gastrectomy is more variable between patients and may not be sustained as long as after RYGB.
For post-surgical patients experiencing weight regain years after bariatric surgery, declining PYY response may be a contributing factor. In these individuals, adding a GLP-1 receptor agonist can restore appetite suppression through a complementary pathway, and the dietary strategies to maximize endogenous PYY production become particularly important for maintaining the hormonal environment that supported initial post-surgical weight loss. The GLP-1 Research Hub discusses GLP-1 therapy for post-bariatric weight regain in more detail.
Binge Eating Disorder and Hedonic Eating
Binge eating disorder (BED) affects approximately 2-3% of the adult population and involves recurring episodes of eating large quantities of food, often rapidly and to the point of discomfort, accompanied by a sense of loss of control. PYY biology is relevant to BED because affected individuals often show blunted PYY responses to meals, meaning their brains receive weaker "stop eating" signals than individuals without BED.
But BED isn't simply a failure of satiety signaling. It involves dysregulation of hedonic (pleasure-based) eating circuits in the brain's reward system. PYY has effects on reward-related brain regions, including the ventral tegmental area and nucleus accumbens, where it can reduce the rewarding value of food cues. This dual action, reducing homeostatic hunger through hypothalamic effects and reducing hedonic eating through reward circuit modulation, makes PYY optimization a theoretically valuable component of BED management.
For individuals with BED, pharmacological appetite control with GLP-1 receptor agonists can reduce binge frequency and severity. Semaglutide has shown particular promise in clinical trials for BED, with significant reductions in binge frequency compared to placebo. Combining pharmacological treatment with PYY-optimizing dietary strategies and, when available, psychological treatment (cognitive-behavioral therapy for BED remains the gold-standard psychological intervention) creates a multi-level approach that addresses the biological, psychological, and behavioral components of the disorder.
Tesofensine, which enhances dopamine and norepinephrine signaling in appetite-regulating circuits, may address the central reward-processing component of binge eating from yet another angle. Its effects on monoamine neurotransmission influence the same reward pathways that PYY modulates, potentially providing complementary benefit. The Comparison Hub offers more on how different weight management compounds target distinct aspects of appetite regulation.
PYY and the Gut Microbiome
The relationship between the gut microbiome and PYY secretion represents one of the most dynamic areas of metabolic research. Different microbial communities produce different profiles and quantities of short-chain fatty acids, which directly stimulate L-cell PYY release. Individuals with microbiome compositions enriched in butyrate-producing bacteria (like Faecalibacterium prausnitzii, Roseburia species, and Eubacterium rectale) tend to have stronger postprandial PYY responses and better metabolic health markers overall.
Antibiotic use, processed food diets, and chronic stress can all shift the microbiome away from SCFA-producing compositions, potentially reducing endogenous PYY output. For individuals on GLP-1 therapy who want to maximize complementary endogenous appetite control, actively supporting SCFA-producing bacteria through prebiotic fiber intake (10-15 grams of inulin, FOS, or resistant starch daily), fermented food consumption, and microbiome-supportive dietary diversity enhances the PYY signaling that works alongside pharmaceutical appetite suppression.
PYY in Adolescents and Young Adults
Childhood and adolescent obesity is one of the most urgent public health challenges, and understanding PYY biology in this population has unique implications. Adolescents generally have more strong PYY responses than adults, which partly explains why some degree of appetite self-regulation occurs during normal development. However, in obese adolescents, PYY responses are blunted similarly to obese adults, suggesting that the PYY dysfunction seen in obesity develops early and may be both a consequence and a perpetuating factor of excess weight.
Interestingly, some research suggests that early-life PYY programming may have lasting effects on appetite regulation throughout adulthood. Infants who are breastfed tend to have different gut hormone profiles than formula-fed infants, with potentially higher PYY responsiveness in later life. The establishment of gut microbiome diversity during early childhood also influences L-cell density and PYY production capacity. These developmental windows highlight how early-life nutrition may program lifelong appetite regulation, a consideration that goes beyond any individual therapeutic intervention.
The treatment of adolescent obesity is complicated by concerns about growth, development, and the long-term effects of pharmacological intervention during a critical developmental window. GLP-1 receptor agonists, specifically semaglutide, have been approved for adolescent use based on clinical trial data, but PYY-focused dietary strategies offer a lower-risk complementary approach. High-protein breakfasts, adequate fiber intake, regular physical activity, and consistent meal timing can optimize PYY signaling without pharmacological intervention, and these habits, if established during adolescence, provide lasting metabolic benefits through adulthood.
The intersection of microbiome science, gut hormone biology, and pharmacological weight management represents a frontier where personalized medicine approaches could significantly improve outcomes. Future testing may allow clinicians to assess a patient's microbiome composition, predict their PYY response potential, and tailor dietary and pharmacological interventions accordingly. For current guidance on nutritional strategies that support both gut health and weight management, the Lifestyle Hub provides evidence-based recommendations, and the FormBlends assessment helps individuals identify the most appropriate combination of dietary, lifestyle, and pharmacological approaches for their specific metabolic profile.
PYY and the Post-Bariatric Surgery Gut Hormone Landscape
Bariatric surgery remains the most effective long-term intervention for severe obesity, and understanding how PYY contributes to its success provides insights that may inform non-surgical approaches to weight management. The dramatic and sustained weight loss following procedures like Roux-en-Y gastric bypass (RYGB) and vertical sleeve gastrectomy (VSG) was historically attributed to mechanical restriction (smaller stomach) and, in the case of RYGB, malabsorption. However, research over the past two decades has fundamentally shifted this understanding toward a gut hormone-centric explanation, with PYY playing a central role.
Following RYGB, fasting and postprandial PYY levels increase dramatically, often reaching 2-3 times pre-surgical levels within weeks of the procedure. This increase occurs because the surgical rearrangement of the intestinal tract delivers partially digested nutrients to the distal ileum (where L-cells that produce PYY are concentrated) much earlier and in higher concentrations than normal anatomy allows. The result is an amplified PYY signal that reduces appetite, slows gastric emptying of the surgical pouch, and shifts food preferences away from high-fat and high-sugar options. VSG produces similar PYY elevations, though typically somewhat less pronounced, likely because the accelerated gastric emptying from the tubularized stomach delivers nutrients to the distal gut faster than a normal stomach would.
What makes the post-bariatric PYY response particularly interesting is its durability. Unlike many other weight loss interventions, where compensatory hunger mechanisms gradually override initial appetite suppression, the elevated PYY levels following bariatric surgery persist for years. Studies with follow-up periods of 5-10 years continue to show elevated PYY responses to meals in post-RYGB patients. This sustained hormonal change, along with similarly persistent elevations in GLP-1, helps explain why bariatric surgery produces durable weight loss where dietary interventions and most pharmacological therapies face the challenge of physiological adaptation and weight regain.
The bariatric surgery model has directly informed efforts to develop PYY-based pharmacological therapies. If elevated PYY is a key mechanism behind bariatric surgery's success, then finding ways to pharmacologically reproduce that PYY elevation, without the surgery, could potentially deliver similar benefits to patients who are not candidates for or who prefer to avoid surgical intervention. Several approaches are being explored: PYY analogs with extended half-lives that could be administered as weekly injections (similar to the GLP-1 receptor agonist model), combination therapies that co-deliver PYY with GLP-1 or other gut hormones to mimic the multi-hormone response seen after bariatric surgery, and oral formulations that stimulate endogenous PYY release from intestinal L-cells.
For patients who have undergone bariatric surgery and are experiencing weight regain years after their procedure, understanding PYY dynamics offers both diagnostic and therapeutic insights. Weight regain after bariatric surgery affects approximately 20-30% of patients and can be devastating psychologically after the effort and risk of surgery. In some cases, weight regain is associated with declining PYY and GLP-1 responses to meals, suggesting that the gut hormone advantage conferred by surgery is partially lost over time, perhaps due to intestinal adaptation or changes in L-cell density. GLP-1 receptor agonists like semaglutide are increasingly used in post-bariatric patients with weight regain, providing pharmacological reinforcement of the gut hormone signal that surgery initially amplified. Combining GLP-1 therapy with dietary strategies that optimize endogenous PYY release, particularly high-protein meals and adequate fiber intake, may provide the most comprehensive approach to managing post-surgical weight regain. The GLP-1 Research Hub covers the evolving evidence on pharmacological approaches to weight management, including post-bariatric applications.
Understanding PYY's role in bariatric surgery also informs the growing interest in endoscopic bariatric procedures, which modify the GI tract less invasively than surgical approaches. Endoscopic sleeve gastroplasty (ESG), intragastric balloons, and other endoscopic techniques produce more modest weight loss than surgery but also generate smaller PYY responses, consistent with the dose-response relationship between anatomical modification and gut hormone change. These intermediate-intensity procedures may be appropriate for patients whose obesity severity doesn't warrant full bariatric surgery but who have not responded adequately to medication alone.
Frequently Asked Questions
References
- Batterham RL, Cowley MA, Small CJ, et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature. 2002;418(6898):650-654. DOI: 10.1038/nature00887.
- Batterham RL, Cohen MA, Ellis SM, et al. Inhibition of food intake in obese subjects by peptide YY3-36. New England Journal of Medicine. 2003;349(10):941-948. DOI: 10.1056/NEJMoa030204.
- Neary NM, Small CJ, Druce MR, et al. Peptide YY3-36 and glucagon-like peptide-17-36 inhibit food intake additively. Endocrinology. 2005;146(12):5120-5127. DOI: 10.1210/en.2005-0237.
- Mentlein R, Dahms P, Grandt D, et al. Proteolytic processing of neuropeptide Y and peptide YY by dipeptidyl peptidase IV. Regulatory Peptides. 1993;49(2):133-144. DOI: 10.1016/0167-0115(93)90435-B.
- Adrian TE, Ferri GL, Bacarese-Hamilton AJ, et al. Human distribution and release of a putative new gut hormone, peptide YY. Gastroenterology. 1985;89(5):1070-1077. DOI: 10.1016/0016-5085(85)90211-2.
- le Roux CW, Batterham RL, Aylwin SJ, et al. Attenuated peptide YY release in obese subjects is associated with reduced satiety. Endocrinology. 2006;147(1):3-8. DOI: 10.1210/en.2005-0972.
- Tatemoto K, Mutt V. Isolation of two novel candidate hormones using a chemical method for finding naturally occurring polypeptides. Nature. 1980;285(5764):417-418. DOI: 10.1038/285417a0.
- Boey D, Lin S, Karl T, et al. Peptide YY ablation in mice leads to the development of hyperinsulinaemia and obesity. Diabetologia. 2006;49(6):1360-1370. DOI: 10.1007/s00125-006-0237-0.
- Boey D, Lin S, Enriquez RF, et al. PYY transgenic mice are protected against diet-induced and genetic obesity. Neuropeptides. 2008;42(1):19-30. DOI: 10.1016/j.npep.2007.11.003.
- le Roux CW, Aylwin SJ, Batterham RL, et al. Gut hormone profiles following bariatric surgery favor an anorectic state, facilitate weight loss, and improve metabolic parameters. Annals of Surgery. 2006;243(1):108-114. DOI: 10.1097/01.sla.0000183349.16877.84.
- Korner J, Bessler M, Cirilo LJ, et al. Effects of Roux-en-Y gastric bypass surgery on fasting and postprandial concentrations of plasma ghrelin, peptide YY, and insulin. Journal of Clinical Endocrinology and Metabolism. 2005;90(1):359-365. DOI: 10.1210/jc.2004-1076.
- Karamanakos SN, Vagenas K, Kalfarentzos F, et al. Weight loss, appetite suppression, and changes in fasting and postprandial ghrelin and peptide-YY levels after Roux-en-Y gastric bypass and sleeve gastrectomy. Annals of Surgery. 2008;247(3):401-407. DOI: 10.1097/SLA.0b013e318156f012.
- Chandarana K, Batterham RL. Shedding pounds after going under the knife: metabolic insights from cutting the gut. Nature Medicine. 2012;18(5):668-669. DOI: 10.1038/nm.2748.
- Schmidt JB, Gregersen NT, Pedersen SD, et al. Effects of PYY3-36 and GLP-1 on energy intake, energy expenditure, and appetite in overweight men. American Journal of Physiology-Endocrinology and Metabolism. 2014;306(11):E1248-E1256. DOI: 10.1152/ajpendo.00569.2013.
- De Silva A, Salem V, Long CJ, et al. The gut hormones PYY 3-36 and GLP-1 7-36 amide reduce food intake and modulate brain activity in appetite centers in humans. Cell Metabolism. 2011;14(5):700-706. DOI: 10.1016/j.cmet.2011.09.010.
- Steinert RE, Feinle-Bisset C, Asarian L, et al. Ghrelin, CCK, GLP-1, and PYY(3-36): secretory controls and physiological roles in eating and glycemia in health, obesity, and after RYGB. Physiological Reviews. 2017;97(1):411-463. DOI: 10.1152/physrev.00031.2014.
- Field BC, Wren AM, Cooke D, et al. Gut hormones as potential new targets for appetite regulation and the treatment of obesity. Drugs. 2008;68(2):147-163. DOI: 10.2165/00003495-200868020-00002.
- Manning S, Batterham RL. The role of gut hormone peptide YY in energy and glucose homeostasis: twelve years on. Annual Review of Physiology. 2014;76:585-608. DOI: 10.1146/annurev-physiol-021113-170404.
- Karra E, Chandarana K, Batterham RL. The role of peptide YY in appetite regulation and obesity. Journal of Physiology. 2009;587(1):19-25. DOI: 10.1113/jphysiol.2008.164269.
- Tan T, Behary P, Tharakan G, et al. The effect of a subcutaneous infusion of GLP-1, OXM, and PYY on energy intake and expenditure in obese volunteers. Journal of Clinical Endocrinology and Metabolism. 2017;102(7):2364-2372. DOI: 10.1210/jc.2017-00469.
- Wulff BS, Catipovic B, Gydesen S, et al. Long-acting PYY3-36 analogue with semaglutide for obesity: from preclinical assessment through randomized clinical studies. Obesity. 2025;33(6):1034-1047. DOI: 10.1002/oby.24329.
- Ruban A, Ashrafian H, Teare JP. The ileocolonic brake to treat obesity: a pilot study. Obesity Surgery. 2018;28(8):2440-2445. DOI: 10.1007/s11695-018-3164-7.
- Sam AH, Gunner DJ, King A, et al. Selective ablation of peptide YY cells in adult mice reveals their role in beta cell survival. Gastroenterology. 2012;143(2):459-468. DOI: 10.1053/j.gastro.2012.04.047.
- Habib AM, Richards P, Cairns LS, et al. Overlap of endocrine hormone expression in the mouse intestine revealed by transcriptional profiling and flow cytometry. Endocrinology. 2012;153(7):3054-3065. DOI: 10.1210/en.2011-2170.
- Degen L, Oesch S, Casanova M, et al. Effect of peptide YY3-36 on food intake in humans. Gastroenterology. 2005;129(5):1430-1436. DOI: 10.1053/j.gastro.2005.09.001.
- Sloth B, Holst JJ, Flint A, et al. Effects of PYY1-36 and PYY3-36 on appetite, energy intake, energy expenditure, glucose and fat metabolism in obese and lean subjects. American Journal of Physiology-Endocrinology and Metabolism. 2007;292(4):E1062-E1068. DOI: 10.1152/ajpendo.00450.2006.
- Stadlbauer U, Woods SC, Langhans W, et al. PYY3-36: beyond food intake. Frontiers in Neuroendocrinology. 2015;38:1-11. DOI: 10.1016/j.yfrne.2014.12.003.
- Goldstone AP, Morgan I, Mercer JG, et al. Effect of leptin on hypothalamic GLP-1 peptide and brain-stem pre-proglucagon mRNA. Biochemical and Biophysical Research Communications. 2000;269(2):331-335. DOI: 10.1006/bbrc.2000.2288.
- Yousseif A, Emmanuel J, Karra E, et al. Differential effects of laparoscopic sleeve gastrectomy and laparoscopic gastric bypass on appetite, circulating acyl-ghrelin, peptide YY3-36 and active GLP-1 levels in non-diabetic humans. Obesity Surgery. 2014;24(2):241-252. DOI: 10.1007/s11695-013-1066-0.
- Nannipieri M, Baldi S, Mari A, et al. Roux-en-Y gastric bypass and sleeve gastrectomy: mechanisms of diabetes remission and role of gut hormones. Journal of Clinical Endocrinology and Metabolism. 2013;98(11):4391-4399. DOI: 10.1210/jc.2013-2538.
- Guida C, Ramracheya R. PYY, a therapeutic option for type 2 diabetes? Clinical Medicine Insights: Endocrinology and Diabetes. 2020;13:1179551419892985. DOI: 10.1177/1179551419892985.
- Dimitriadis GK, Randeva MS, Kaltsas G. Peptide YY in type 2 diabetes: a complementary gut hormone with therapeutic potential beyond GLP-1. Nutrients. 2025;17(21):3468. DOI: 10.3390/nu17213468.
- Svendsen B, Pedersen J, Albrechtsen NJ, et al. An analysis of cosecretion and coexpression of gut hormones from male rat proximal and distal small intestine. Endocrinology. 2015;156(3):847-857. DOI: 10.1210/en.2014-1710.
- Psichas A, Sleeth ML, Murphy KG, et al. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. International Journal of Obesity. 2015;39(3):424-429. DOI: 10.1038/ijo.2014.153.
- Ueda SY, Yoshikawa T, Katsura Y, et al. Changes in gut hormone levels and negative energy balance during aerobic exercise in obese young males. Journal of Endocrinology. 2009;201(1):151-159. DOI: 10.1677/JOE-08-0500.
- Pannacciulli N, Bunt JC, Kober SM, et al. Higher fasting plasma concentrations of glucagon-like peptide 1 are associated with higher resting energy expenditure and lower body weight. Journal of Clinical Endocrinology and Metabolism. 2006;91(10):3935-3940. DOI: 10.1210/jc.2006-0868.