Executive Summary
LL-37 is the only cathelicidin antimicrobial peptide produced in the human body. It serves as a front-line weapon against bacteria, viruses, fungi, and parasites while simultaneously orchestrating immune responses, promoting wound repair, and breaking apart stubborn biofilms. For researchers, clinicians, and anyone interested in the intersection of peptide science and infection defense, LL-37 represents one of the most versatile molecules in the human immune arsenal.
Key Takeaways
- LL-37 is the sole human cathelicidin - a 37-amino-acid peptide with broad-spectrum antimicrobial activity against bacteria, viruses, fungi, and biofilms
- Vitamin D directly regulates LL-37 production through a vitamin D response element in the CAMP gene promoter
- Beyond killing pathogens, LL-37 modulates innate and adaptive immune responses, promotes wound healing, and enhances angiogenesis
- Clinical trials have demonstrated safety and efficacy in hard-to-heal venous leg ulcers with topical LL-37 application
- LL-37 disrupts bacterial biofilms through multiple mechanisms, including interference with initial attachment and disruption of quorum sensing
Your body makes LL-37 every day. Neutrophils store it in granules, ready to release at the first sign of microbial invasion. Epithelial cells lining your skin, lungs, gut, and urinary tract produce it continuously. And vitamin D - the so-called sunshine vitamin - directly controls how much your cells manufacture. This single connection between sun exposure, vitamin D status, and antimicrobial peptide production helps explain why respiratory infections spike in winter months when vitamin D levels drop.
But LL-37 does far more than kill pathogens. It recruits immune cells to sites of infection. It tells macrophages when to ramp up their inflammatory response and when to dial it back. It stimulates the growth of new blood vessels and accelerates wound closure. It even disrupts the protective biofilm shields that bacteria construct to resist antibiotics. In an era of rising antibiotic resistance, these properties have made LL-37 one of the most studied peptides in infectious disease research.
This report examines the molecular biology of LL-37, its mechanisms of antimicrobial action, its roles in immune modulation and wound healing, the vitamin D connection, and the current state of clinical research. We'll also explore how LL-37 compares to and works alongside other immune-supportive peptides like Thymosin Alpha-1 and KPV, practical dosing considerations from available research, and safety data.
Key Takeaways
- LL-37 is the sole human cathelicidin - a 37-amino-acid peptide with broad-spectrum antimicrobial activity against bacteria, viruses, fungi, and biofilms
- Vitamin D directly regulates LL-37 production through a vitamin D response element in the CAMP gene promoter
- Beyond killing pathogens, LL-37 modulates innate and adaptive immune responses, promotes wound healing, and enhances angiogenesis
- Clinical trials have demonstrated safety and efficacy in hard-to-heal venous leg ulcers with topical LL-37 application
- LL-37 disrupts bacterial biofilms through multiple mechanisms, including interference with initial attachment and disruption of quorum sensing
- Research suggests anti-tumor properties in certain cancer models, adding another dimension to this peptide's therapeutic potential

Human Antimicrobial Peptide Biology
The Cathelicidin Gene and Protein Processing
The story of LL-37 begins with a single gene: CAMP (cathelicidin antimicrobial peptide), located on chromosome 3 at position 3p21.31. This gene encodes a precursor protein called hCAP18 (human cationic antimicrobial protein 18 kDa), which consists of three distinct regions: a signal peptide that directs it to the correct cellular compartment, a cathelin-like domain that keeps the peptide inactive during storage, and the C-terminal LL-37 peptide itself (Zanetti, 2004).
Think of hCAP18 as a loaded weapon with the safety on. The cathelin-like domain acts as that safety mechanism, preventing LL-37 from damaging the cells that produce and store it. When neutrophils degranulate in response to infection, the enzyme proteinase 3 cleaves hCAP18 at a specific site, releasing the active 37-amino-acid LL-37 peptide into the extracellular environment (Sorensen et al., 2001). Epithelial cells use a different set of enzymes - kallikreins, particularly kallikrein 5 and kallikrein 7 - to process the precursor, which allows tissue-specific fine-tuning of antimicrobial peptide production.
Neutrophils are by far the richest source of LL-37. Each neutrophil stores approximately 630 ng of hCAP18 in its specific granules, and with billions of neutrophils circulating at any given time, the body maintains a massive reservoir of this antimicrobial peptide. But neutrophils aren't the only producers. Epithelial cells of the skin, respiratory tract, gastrointestinal tract, and urogenital system all express CAMP, as do monocytes, macrophages, mast cells, natural killer cells, and certain lymphocyte subsets (Durr et al., 2006).
LL-37 Structure: An Amphipathic Alpha-Helix
The name LL-37 tells you two things about this peptide's structure: it begins with two leucine residues (LL) and it contains 37 amino acids. Its full amino acid sequence is LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES. At physiological pH, the peptide carries a net positive charge of +6, which is critical for its interaction with negatively charged bacterial membranes.
What makes LL-37 so effective as an antimicrobial agent is its amphipathic alpha-helical structure. When LL-37 encounters a membrane or membrane-like environment, it folds into an alpha-helix where all the positively charged and hydrophilic residues line up on one face, while the hydrophobic residues cluster on the opposite face. This creates a molecular structure with one side that loves water and another side that loves lipids - the perfect configuration for inserting into and disrupting cellular membranes (Wang, 2008).
NMR studies have revealed that LL-37 in solution consists of an N-terminal helical domain (residues 2-31) and a disordered C-terminal tail (residues 32-37). The helical region can be further divided into a hydrophobic N-terminal helix and an amphipathic C-terminal helix connected by a bend around residues 14-16. This structural flexibility allows LL-37 to adapt its conformation depending on the membrane composition it encounters, which partially explains its remarkably broad antimicrobial spectrum (Porcelli et al., 2008).
At higher concentrations, LL-37 can form oligomers. Crystallographic studies show that four LL-37 molecules can come together to create a channel-like structure capable of forming transmembrane pores. This oligomerization is concentration-dependent and influenced by the lipid composition of the target membrane, meaning LL-37's killing mechanism can shift between different modes depending on local conditions (Sancho-Vaello et al., 2020).
Research Note
The amphipathic structure of LL-37 is shared by many other antimicrobial peptides across species, suggesting this molecular architecture evolved independently multiple times as an optimal design for membrane disruption. However, LL-37's specific sequence and structural features give it immunomodulatory properties that go well beyond simple membrane lysis, distinguishing it from many other antimicrobial peptides.
Tissue Distribution and Expression Patterns
LL-37 shows up wherever the body interfaces with the outside world. In healthy skin, keratinocytes produce constitutive low levels that ramp up dramatically during infection or wounding. The respiratory epithelium maintains baseline LL-37 expression that increases during upper and lower respiratory tract infections. In the gastrointestinal tract, colonic epithelial cells and Paneth cells of the small intestine both contribute to luminal LL-37 concentrations that help shape the gut microbiome.
Circulating concentrations of LL-37 in healthy adults typically range from 1 to 5 micrograms per milliliter of plasma. At sites of active infection or inflammation, local concentrations can spike to 20 micrograms per milliliter or higher. These high local concentrations are sufficient to kill most susceptible pathogens directly, while even sub-bactericidal concentrations exert significant immunomodulatory effects.
Expression patterns vary significantly between individuals and are influenced by multiple factors. Vitamin D status is the most well-characterized regulator (discussed in detail in a later section), but butyrate from gut bacterial fermentation, infection-related cytokines like IL-1beta and IL-6, and toll-like receptor activation all influence CAMP gene transcription. This multi-input regulation allows the body to fine-tune LL-37 production based on the specific threats it faces.
Researchers interested in immune defense peptides may also want to explore Thymosin Alpha-1, which works through complementary pathways to enhance innate immune surveillance, or visit the Peptide Research Hub for a broader overview of immune-supportive compounds.

Membrane Disruption Mechanism
Step 1: Electrostatic Attraction and Initial Binding
The killing starts with charge. Bacterial membranes carry a strong negative charge on their outer surface due to the presence of lipopolysaccharide (LPS) in Gram-negative bacteria and lipoteichoic acid (LTA) in Gram-positive species. LL-37's net positive charge of +6 creates an electrostatic attraction that draws the peptide toward bacterial surfaces while largely sparing human cells, whose outer membranes are composed predominantly of neutral (zwitterionic) phospholipids like phosphatidylcholine and sphingomyelin.
This charge-based selectivity is the primary reason why LL-37 preferentially targets microbial cells over host cells. Cholesterol in mammalian membranes provides additional protection by reducing the ability of LL-37 to insert into and disrupt the lipid bilayer. However, at very high concentrations, LL-37 can overcome this selectivity and damage host cells - a characteristic that becomes relevant in certain inflammatory conditions and that limits maximum therapeutic dosing.
Step 2: Membrane Insertion and Disruption
Once LL-37 reaches the bacterial surface, it follows a multi-step process of membrane disruption. For Gram-negative bacteria, the peptide must first cross the outer membrane by interacting with and displacing divalent cations (Mg2+ and Ca2+) that normally stabilize LPS packing. This destabilizes the outer membrane and grants access to the inner (cytoplasmic) membrane, which is the primary target.
Research has identified two distinct modes of membrane disruption that LL-37 employs, depending on the lipid composition of the target membrane. In membranes rich in unsaturated phospholipids, LL-37 forms classical transmembrane pores - organized channels that allow the rapid efflux of intracellular contents. In membranes containing primarily saturated phospholipids, the peptide instead causes general membrane thinning and disordering without forming discrete pores (Xhindoli et al., 2016).
The pore-forming mechanism involves LL-37 oligomerization at the membrane surface, followed by insertion of the oligomeric complex through the lipid bilayer. Crystallographic and simulation studies suggest that four LL-37 molecules come together to create a toroidal pore approximately 2-3 nanometers in diameter - large enough to allow passage of ions, small molecules, and even some proteins. At high peptide-to-lipid ratios, LL-37 can also induce the formation of membrane-spanning nanofibers that create even larger disruptions.
Step 3: Intracellular Targets
Membrane disruption is not the whole story. At sub-lytic concentrations - levels too low to cause outright membrane destruction - LL-37 can still kill bacteria by reaching intracellular targets. Once inside the cell, the peptide can bind to bacterial DNA, inhibiting replication and transcription. It can also interfere with protein synthesis by binding to ribosomes, disrupt cell wall synthesis by inhibiting enzymes in the peptidoglycan biosynthetic pathway, and trigger oxidative stress by disrupting electron transport chain components.
This multi-target mechanism of action is one reason why bacteria have great difficulty developing resistance to LL-37. Unlike conventional antibiotics that typically hit a single molecular target (which can be altered by a single mutation), LL-37 attacks bacteria on multiple fronts simultaneously. A bacterium would need to fundamentally redesign its membrane composition, alter its DNA-binding properties, and modify multiple intracellular targets all at once - an evolutionary challenge of enormous proportions.
Clinical Significance
The multi-target mechanism of LL-37 makes it particularly valuable in the context of antibiotic-resistant infections. While MRSA, VRE, and multidrug-resistant Gram-negative organisms have accumulated resistance mechanisms against multiple antibiotic classes, these resistance mechanisms are largely ineffective against LL-37's membrane-disruption approach. Research has confirmed that LL-37 retains full activity against clinical isolates of MRSA and other drug-resistant pathogens.
Speed of Killing Action
One of LL-37's most impressive characteristics is its speed. Time-kill studies have demonstrated that LL-37 can achieve greater than 3-log reductions (99.9% killing) of S. aureus within 5 minutes of contact. For E. coli, significant killing occurs within 15-30 minutes at concentrations above the minimum inhibitory concentration (MIC). This rapid action contrasts sharply with conventional antibiotics, many of which require hours to days to achieve comparable bactericidal effects.
The speed of LL-37's action makes biological sense. When neutrophils arrive at a site of infection and degranulate, releasing stored LL-37 along with other antimicrobial factors, the immune system needs rapid pathogen killing to prevent bacterial dissemination. A slow-acting antimicrobial would be of limited value in this context. The near-instantaneous membrane disruption caused by LL-37 provides exactly the kind of rapid, decisive antimicrobial action that the innate immune system requires.
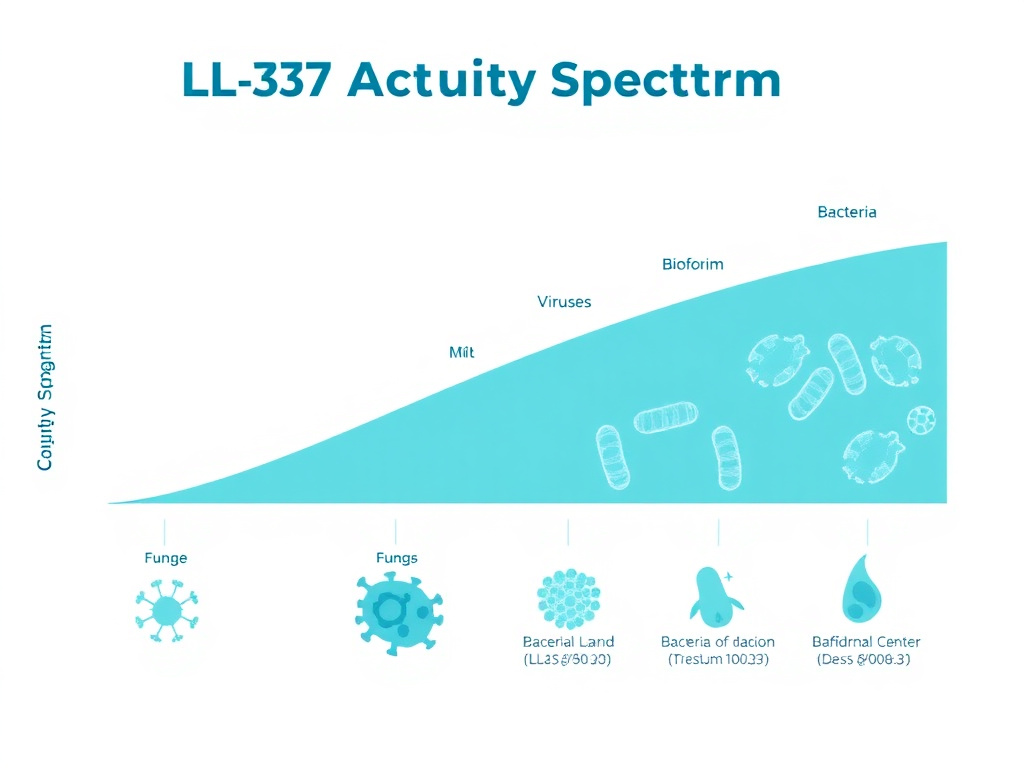
Antimicrobial Spectrum
Antibacterial Activity
LL-37 kills a remarkably broad range of bacterial species. Published research documents direct antimicrobial activity against over 38 different bacterial species, spanning both Gram-positive and Gram-negative categories. The minimum inhibitory concentrations (MICs) vary depending on the specific pathogen, the growth conditions, and the assay methodology, but generally fall in the range of 1-32 micrograms per milliliter.
LL-37 MIC Values Against Common Pathogens
MIC values represent minimum concentrations needed to inhibit bacterial growth in standard in vitro assays. Lower values indicate greater potency.
Among Gram-positive bacteria, LL-37 shows strong activity against Staphylococcus aureus (including methicillin-resistant strains), Streptococcus pyogenes, Streptococcus pneumoniae, Enterococcus faecalis, and Listeria monocytogenes. For Gram-negative species, susceptible organisms include Escherichia coli, Pseudomonas aeruginosa, Klebsiella pneumoniae, Acinetobacter baumannii, Salmonella typhimurium, and Helicobacter pylori.
The antibacterial activity of LL-37 is influenced by environmental conditions. Physiological salt concentrations (150 mM NaCl) can reduce LL-37's potency against certain species by 2-4 fold, as sodium ions compete with the peptide for binding sites on the bacterial surface. Divalent cations like Mg2+ and Ca2+ have an even stronger antagonistic effect. Serum components, particularly high-density lipoprotein (HDL) and apolipoprotein A-I, can bind LL-37 and sequester it from its antimicrobial function. These factors need to be considered when translating in vitro MIC data to in vivo efficacy predictions.
| Pathogen | MIC Range (microg/mL) | Activity Level | Clinical Relevance |
|---|---|---|---|
| Staphylococcus aureus (MSSA) | 2-8 | Strong | Skin/soft tissue infections |
| MRSA | 2-8 | Strong | Hospital-acquired infections |
| Escherichia coli | 1-4 | Very Strong | UTIs, GI infections |
| Pseudomonas aeruginosa | 4-16 | Moderate | Wound infections, CF |
| Klebsiella pneumoniae | 4-16 | Moderate | Pneumonia, UTIs |
| Streptococcus pyogenes | 1-4 | Very Strong | Pharyngitis, cellulitis |
| Enterococcus faecalis | 8-32 | Moderate | Endocarditis, UTIs |
| Acinetobacter baumannii | 4-16 | Moderate | Wound/ICU infections |
| Helicobacter pylori | 2-8 | Strong | Gastric ulcers |
| Candida albicans | 8-32 | Moderate | Mucosal candidiasis |
Antiviral Activity
LL-37's antiviral properties represent one of the most exciting areas of current research. The peptide has demonstrated direct activity against at least 16 different viral species, including both enveloped and non-enveloped viruses. The mechanisms of antiviral action differ from antibacterial killing and include direct disruption of viral envelopes, blocking of viral entry into host cells, and modulation of host antiviral immune responses.
Influenza virus has been the most thoroughly studied viral target. Research by Barlow et al. (2011) demonstrated that LL-37 reduced lung viral titers in mice by approximately 70-80% at 3 days post-infection - a level of reduction comparable to that achieved by the prescription antiviral zanamivir (Tamiflu). Electron microscopy studies revealed that LL-37 directly disrupts the influenza viral membrane, causing fragmentation and loss of structural integrity. The peptide also interferes with viral hemagglutinin-mediated binding to host cell sialic acid residues, blocking the initial attachment step.
Against respiratory syncytial virus (RSV), LL-37 inhibits viral replication in polarized airway epithelial cells at concentrations achievable in the respiratory tract during inflammation. The peptide acts through direct virucidal activity and also enhances epithelial cell antiviral defenses by stimulating interferon-stimulated gene expression (Currie et al., 2016).
LL-37 also shows activity against herpes simplex virus (HSV-1 and HSV-2), human immunodeficiency virus (HIV-1), vaccinia virus, dengue virus, and Zika virus. For enveloped viruses, the primary mechanism appears to be direct disruption of the lipid envelope - essentially the same membrane-disruption mechanism used against bacteria. For some non-enveloped viruses, the mechanism involves blocking viral attachment to host cell receptors or interfering with intracellular viral replication steps.
Antifungal Activity
The antifungal properties of LL-37 extend to at least 16 fungal species. Candida albicans, the most common human fungal pathogen, is susceptible to LL-37 at MIC values of 8-32 micrograms per milliliter. Other susceptible Candida species include C. krusei, C. tropicalis, and C. parapsilosis. LL-37 also shows activity against Aspergillus fumigatus, dermatophytes like Trichophyton rubrum, and the opportunistic mold Fusarium.
The antifungal mechanism involves disruption of the fungal cell membrane, similar to the antibacterial mechanism but requiring higher peptide concentrations due to the different membrane composition of fungal cells. The presence of ergosterol (rather than cholesterol) in fungal membranes and the thick fungal cell wall both contribute to the relatively higher MICs seen against fungi compared to bacteria.
For readers interested in antifungal peptide applications, the Science & Research section at FormBlends provides additional resources on peptides with antimicrobial properties.
Wound Healing Enhancement
Cellular Mechanisms of Wound Repair
LL-37 accelerates wound healing through multiple interconnected mechanisms that go well beyond simply killing bacteria at the wound site. The peptide directly stimulates the proliferation and migration of keratinocytes, fibroblasts, and endothelial cells - the three cell types most critical for wound closure and tissue regeneration. It does this through activation of several receptor systems, including the epidermal growth factor receptor (EGFR), formyl peptide receptor-like 1 (FPRL-1), and purinergic receptor P2X7.
The EGFR pathway is particularly interesting. LL-37 acts as an indirect EGFR ligand through a process called transactivation. The peptide activates metalloproteinases on the cell surface that cleave membrane-bound EGFR ligands (like HB-EGF and amphiregulin), releasing them to bind and activate EGFR. This triggers downstream Ras/MAPK and PI3K/Akt signaling cascades that drive cell proliferation, migration, and survival (Heilborn et al., 2003).
Angiogenesis - the formation of new blood vessels - is another critical component of wound healing that LL-37 promotes. The peptide stimulates endothelial cell proliferation and tube formation through FPRL-1-dependent signaling. In animal models, wounds treated with LL-37 show significantly increased vascular density compared to untreated controls, which translates to better nutrient and oxygen delivery to the healing tissue.
LL-37 also influences the inflammatory phase of wound healing. In the early stages after injury, the peptide promotes a pro-inflammatory response that helps clear debris and kill contaminating bacteria. As healing progresses, LL-37 shifts toward anti-inflammatory signaling, reducing excessive neutrophil infiltration and promoting the transition from inflammatory to proliferative wound healing phases. This dual regulatory capacity makes LL-37 particularly effective in chronic wounds, where persistent inflammation often stalls the healing process.
Application in Chronic Wounds
Chronic wounds - including venous leg ulcers, diabetic foot ulcers, and pressure injuries - represent a massive clinical burden, affecting approximately 2.5% of the U.S. population and costing the healthcare system over $25 billion annually. Many chronic wounds are colonized or infected with biofilm-forming bacteria, particularly S. aureus and P. aeruginosa, which resist conventional antibiotic therapy and perpetuate the inflammatory state that prevents healing.
LL-37 addresses chronic wound pathology on multiple fronts simultaneously. It kills planktonic bacteria, disrupts established biofilms (detailed in a later section), reduces excessive inflammation, stimulates epithelial cell migration, and promotes angiogenesis. This combination of antimicrobial, anti-inflammatory, and pro-regenerative activities makes it uniquely suited for chronic wound therapy.
A landmark clinical study by Gronberg et al. (2014) evaluated topical LL-37 treatment in patients with hard-to-heal venous leg ulcers. This randomized, placebo-controlled, double-blind, phase I/II trial enrolled 34 patients and demonstrated that LL-37 treatment was safe and effective, significantly accelerating wound healing compared to placebo. No safety concerns were identified regarding local or systemic adverse events. The trial used a topical formulation applied directly to the wound bed twice weekly for 4 weeks.
The wound healing properties of LL-37 complement those of other peptides studied for tissue repair. BPC-157 is another peptide extensively researched for its wound healing and tissue repair capabilities, working through partially overlapping but distinct mechanisms. The combination of antimicrobial-driven wound healing (LL-37) with tissue-protective and angiogenic mechanisms (BPC-157) represents an area of growing research interest. For comprehensive information on BPC-157's mechanisms, see the BPC-157 research report.
LL-37-Derived Wound Therapies
Researchers have developed several LL-37-derived peptides that maintain or improve upon the parent molecule's wound healing properties while addressing some of its limitations. P60.4Ac, a truncated analog, retains antimicrobial and wound healing activity while showing improved stability and reduced cytotoxicity. LL-37-loaded hydrogels and nanoparticle formulations have been designed to provide sustained release at wound sites, extending the duration of therapeutic activity.
In thermally wounded human skin equivalents (a laboratory model of burn injuries), LL-37-derived peptides P60.4Ac and P10 eradicated multidrug-resistant S. aureus while promoting re-epithelialization. These results suggest that LL-37-based therapies could be particularly valuable for burn wound management, where infection control and rapid wound closure are both critical priorities (de Breij et al., 2015).
Clinical Evidence Summary
The only completed clinical trial of LL-37 for wound healing (Gronberg et al., 2014) demonstrated safety and efficacy in venous leg ulcers. Additional clinical trials are planned or underway for other wound types. While the existing clinical evidence is limited to a single phase I/II trial, the strong preclinical data across multiple wound models supports continued development.
Vitamin D and LL-37 Expression
The Vitamin D Response Element
One of the most clinically significant discoveries about LL-37 is that its production is directly controlled by vitamin D. The CAMP gene promoter contains a vitamin D response element (VDRE) that binds the vitamin D receptor (VDR) in complex with retinoid X receptor (RXR). When 1,25-dihydroxyvitamin D3 (the active hormonal form of vitamin D, also called calcitriol) binds to this receptor complex, it directly upregulates CAMP gene transcription, increasing LL-37 production (Wang et al., 2004).
This discovery, first reported by Wang and colleagues in 2004, fundamentally changed our understanding of vitamin D's role in immune defense. It provided a molecular mechanism explaining why vitamin D deficiency is associated with increased susceptibility to infections, and it transformed cathelicidin from a curiosity of innate immunity into a central node connecting nutritional status to antimicrobial defense.
The vitamin D-cathelicidin axis is specific to humans and other primates. The VDRE in the CAMP gene promoter is present in primate genomes but absent in mice and other rodents, which is why early murine studies missed this connection entirely. This species difference has important implications for research methodology - mouse models cannot recapitulate the vitamin D-LL-37 regulatory axis, limiting their utility for studying this aspect of human immunity.
Clinical Correlation: Vitamin D Levels and LL-37
Multiple clinical studies have confirmed a positive correlation between circulating 25-hydroxyvitamin D levels and plasma hCAP18/LL-37 concentrations. Dixon et al. (2012) found a significant positive correlation in healthy adults, with the strongest correlation observed in subjects with 25(OH)D concentrations below 32 ng/mL. Above this threshold, the correlation plateaued, suggesting that there may be a vitamin D saturation point for LL-37 induction.
This finding has practical implications. It suggests that vitamin D supplementation is most likely to boost LL-37 levels in individuals who are deficient or insufficient (below 30 ng/mL), while those with replete levels may see diminishing returns from further supplementation. Given that an estimated 42% of American adults are vitamin D deficient and 82% of Black Americans have suboptimal levels, the potential population-level impact of optimizing vitamin D status on antimicrobial defense is substantial.
Several intervention studies have directly tested whether vitamin D supplementation increases LL-37 levels. In tuberculosis contacts, high-dose vitamin D supplementation significantly increased circulating LL-37 concentrations and enhanced antimycobacterial activity of blood cells ex vivo. In neonates, vitamin D3 administration induced expression of CAMP and increased LL-37 in blood cells (Mandic Havelka et al., 2010). Topical application of the vitamin D analogue calcipotriol enhanced hCAP18/LL-37 upregulation during wounding in human skin in vivo (Schauber et al., 2009).
Cell-Type Specific Responses to Vitamin D
Different immune cell types show varying responses to vitamin D-mediated CAMP induction. Gombart et al. (2005) found that in peripheral blood-derived cells, macrophages showed the highest levels of hCAP18 expression following treatment with 1,25D3, while GM-CSF-derived dendritic cells and osteoclasts showed comparatively lower levels. Neutrophils, which have the highest baseline hCAP18 expression among circulating immune cells, showed more modest vitamin D-dependent upregulation, possibly because their expression is already near maximal under normal conditions.
In epithelial cells, the vitamin D response varies by tissue. Respiratory epithelial cells show strong CAMP upregulation in response to vitamin D, which may partially explain the observed association between vitamin D deficiency and respiratory infections. Intestinal epithelial cells also respond to vitamin D with increased LL-37 production, linking vitamin D status to gut mucosal defense. Skin keratinocytes upregulate LL-37 in response to both vitamin D and injury, creating a coordinated wound defense mechanism.
Practical Implication
Maintaining adequate vitamin D levels (at least 30 ng/mL, with many experts recommending 40-60 ng/mL) is one of the simplest and most evidence-based strategies for supporting endogenous LL-37 production. This represents a low-cost, low-risk approach to enhancing natural antimicrobial defenses that can complement more targeted peptide-based interventions.
Immune Modulation
Chemotaxis and Immune Cell Recruitment
LL-37 is a potent chemoattractant for multiple immune cell types. It recruits monocytes, T cells, neutrophils, and mast cells to sites of infection through direct activation of the formyl peptide receptor-like 1 (FPRL-1, now designated FPR2/ALX). This chemotactic activity operates at nanomolar concentrations - well below the levels needed for direct antimicrobial killing - meaning that even trace amounts of LL-37 released during the earliest stages of infection can begin rallying immune cell reinforcements.
The recruitment hierarchy follows a logical pattern. Neutrophils, which respond fastest and carry the most pre-formed LL-37 in their granules, are among the first responders attracted by initial LL-37 release. As they arrive and degranulate, they release additional LL-37 that amplifies the chemotactic signal, recruiting monocytes that will differentiate into macrophages for sustained pathogen clearance and tissue repair. T cells arrive later, guided in part by LL-37's chemotactic gradient, to provide adaptive immune support.
Beyond simple chemotaxis, LL-37 also stimulates mast cell degranulation, releasing histamine and other mediators that increase local blood flow and vascular permeability. This facilitates the extravasation (tissue entry) of additional immune cells from the bloodstream. The peptide also induces mast cell production of cytokines including IL-4, IL-5, and IL-6, further shaping the local immune environment.
Cytokine Modulation: Dual Pro- and Anti-Inflammatory Effects
One of LL-37's most remarkable properties is its ability to both promote and suppress inflammation depending on the context. This dual regulatory capacity prevents both inadequate immune responses (which could allow infections to spread) and excessive inflammation (which causes tissue damage).
In the pro-inflammatory direction, LL-37 stimulates IL-8 secretion from epithelial cells, monocytes, and other cell types, promoting further neutrophil recruitment. It enhances flagellin-induced IL-8 production in keratinocytes through P2X7 receptor signaling and Src/Akt pathway activation. It promotes the secretion of IL-6, a cytokine with both pro- and anti-inflammatory properties, from bronchial epithelial cells via the PI3K/p38 pathway. And it stimulates dendritic cell maturation, enhancing antigen presentation and bridging innate and adaptive immunity (Bowdish et al., 2005).
In the anti-inflammatory direction, LL-37 neutralizes bacterial endotoxin (LPS), preventing the excessive inflammatory response that can lead to septic shock. It suppresses LPS-induced TNF-alpha and IL-6 production from macrophages, reducing the risk of cytokine storm. In neutrophils, LL-37 decreases the release of multiple pro-inflammatory cytokines while simultaneously enhancing bacterial phagocytosis and reactive oxygen species production - essentially making neutrophils better killers with less collateral inflammation (Zheng et al., 2007).
The signaling mechanisms underlying these opposing effects have been partially characterized. In monocytes, LL-37's anti-inflammatory effects are mediated through inhibition of STAT1-independent signaling, involving both the p65 subunit of NF-kappaB and p38 MAPK. The balance between pro- and anti-inflammatory signaling appears to depend on the local concentration of LL-37, the presence or absence of microbial products, and the activation state of the target cells.
This immune-modulatory profile makes LL-37 conceptually similar to other immune peptides. KPV, an alpha-MSH fragment, also exhibits powerful anti-inflammatory properties particularly relevant to gut and skin inflammation. Thymosin Alpha-1 modulates immune function at a different level, primarily enhancing T cell and dendritic cell function. The Thymosin Alpha-1 research report provides detailed coverage of that peptide's immune-modulating mechanisms.
Toll-Like Receptor Interactions
LL-37 modulates Toll-like receptor (TLR) signaling in complex and context-dependent ways. It can bind extracellular double-stranded DNA and RNA, forming complexes that are internalized by immune cells and delivered to endosomal TLRs (specifically TLR3, TLR7, TLR8, and TLR9). This property has important implications for both antimicrobial defense and autoimmune disease.
In the context of infection, LL-37's ability to deliver microbial nucleic acids to endosomal TLRs enhances the detection of intracellular pathogens. By complexing with bacterial or viral DNA/RNA and facilitating their uptake, LL-37 essentially acts as a delivery vehicle that amplifies pattern recognition receptor signaling. This enhances type I interferon production and activates plasmacytoid dendritic cells, strengthening antiviral defenses.
However, this same property can become problematic in autoimmune conditions. In psoriasis, LL-37 forms complexes with self-DNA released from damaged keratinocytes and delivers it to TLR9 in plasmacytoid dendritic cells, triggering an inappropriate type I interferon response that perpetuates skin inflammation. This mechanism has been identified as a key driver of psoriatic pathology and illustrates how LL-37's immune functions, while generally protective, can contribute to disease under certain conditions (Lande et al., 2007).
Effects on Natural Killer Cells and Adaptive Immunity
LL-37 influences adaptive immunity both directly and through its effects on dendritic cells. The peptide enhances dendritic cell differentiation from monocytes, increases antigen uptake and processing, upregulates co-stimulatory molecules (CD80, CD86), and promotes Th1-polarized T cell responses. These effects position LL-37 as a bridge between innate detection of pathogens and the development of targeted adaptive immune responses.
Natural killer (NK) cells are also affected by LL-37. The peptide enhances NK cell cytotoxicity against tumor cells and virus-infected cells, potentially contributing to both anti-infection and anti-tumor immune surveillance. This effect appears to be mediated through FPRL-1 signaling and is enhanced in the presence of IL-2 or IL-12.
Biofilm Disruption
The Biofilm Problem
Bacterial biofilms are communities of microorganisms encased in a self-produced matrix of extracellular polymeric substances (EPS) - a mixture of polysaccharides, proteins, lipids, and extracellular DNA. Biofilms are clinically significant because bacteria within them are 100 to 1,000 times more resistant to conventional antibiotics than their free-floating (planktonic) counterparts. An estimated 80% of chronic infections involve biofilm-forming organisms, including chronic wound infections, implant-associated infections, chronic sinusitis, chronic urinary tract infections, and cystic fibrosis lung infections.
The medical device industry faces particular challenges with biofilms. Catheters, joint prostheses, heart valves, and dental implants all provide surfaces where biofilms readily form. Once established, these biofilms are extremely difficult to eradicate without removing the device itself. LL-37's anti-biofilm properties address this critical clinical need through mechanisms that are distinct from its direct antimicrobial (planktonic) activity.
Anti-Biofilm Mechanisms
LL-37 combats biofilms through at least four distinct mechanisms, and understanding each one reveals why this peptide is so effective against these resistant communities.
Prevention of Initial Attachment: At sub-inhibitory concentrations (below the MIC for planktonic bacteria), LL-37 reduces bacterial adhesion to surfaces by altering the expression of genes involved in surface attachment. In P. aeruginosa, sub-MIC concentrations of LL-37 downregulate genes required for type IV pili assembly and flagellar motility, reducing the bacterium's ability to approach and adhere to surfaces in the first place (Overhage et al., 2008).
Stimulation of Surface Motility: LL-37 promotes twitching motility in P. aeruginosa, a form of surface movement powered by type IV pili extension and retraction. Increased twitching motility prevents the dense cell-cell contacts needed for biofilm maturation, essentially keeping bacteria in a dispersed, planktonic-like state where they remain susceptible to antimicrobial agents and immune cell killing.
Quorum Sensing Interference: Bacteria coordinate biofilm formation through chemical signaling molecules called quorum sensing (QS) signals. LL-37 disrupts QS in multiple species by interfering with signal molecule production, degradation, or receptor binding. This prevents the coordinated gene expression changes that drive the transition from planktonic growth to biofilm formation.
Disruption of Established Biofilms: Against pre-formed biofilms, LL-37 can penetrate the EPS matrix and kill embedded bacteria. Unlike many antibiotics that are excluded from the biofilm interior by the EPS barrier, LL-37's cationic character allows it to interact with negatively charged EPS components and migrate through the matrix. Once inside, the peptide disrupts bacterial membranes and causes biofilm detachment and dispersal.
Research Data on Biofilm Efficacy
Overhage et al. (2008) demonstrated that LL-37 at concentrations as low as 0.5 micrograms per milliliter (well below the MIC of 64 micrograms per milliliter against planktonic P. aeruginosa in their assay) significantly inhibited P. aeruginosa biofilm formation. This sub-MIC anti-biofilm activity is clinically meaningful because it means LL-37 can prevent biofilm formation at concentrations far below those needed for direct bacterial killing.
Against pre-formed S. aureus biofilms, Noore et al. (2013) showed that LL-37 achieved rapid bactericidal activity with greater than 3-log reduction within 5 minutes of treatment. The speed of action against biofilm-embedded bacteria was comparable to that against planktonic cells, suggesting that LL-37 is not significantly hindered by the EPS matrix barrier that renders many conventional antibiotics ineffective.
Combination studies have explored LL-37 paired with conventional antibiotics for enhanced anti-biofilm activity. Dosler and Karaaslan (2014) found that LL-37 combined with polymyxin B showed significant prevention of biofilm formation and eradication of pre-grown E. coli and P. aeruginosa biofilms. The combination was more effective than either agent alone, suggesting complementary mechanisms of action.
| Biofilm Species | LL-37 Concentration | Effect | Study |
|---|---|---|---|
| P. aeruginosa (prevention) | 0.5 microg/mL | Significant inhibition of formation | Overhage et al., 2008 |
| S. aureus (pre-formed) | 16-32 microg/mL | >3 log kill in 5 min | Noore et al., 2013 |
| E. coli (pre-formed) | 8-16 microg/mL | Significant eradication | Dosler & Karaaslan, 2014 |
| MRSA (pre-formed) | 16-64 microg/mL | Disruption and killing | Dean et al., 2011 |
| C. albicans (pre-formed) | 32-64 microg/mL | Biofilm mass reduction | Tsai et al., 2011 |
Biofilm Relevance
The anti-biofilm properties of LL-37 are particularly significant because they operate at concentrations below the MIC for planktonic killing. This means that even at tissue concentrations too low to kill free-floating bacteria directly, LL-37 can still prevent biofilm formation and maintain bacterial susceptibility to other immune defenses and antibiotic therapies. This sub-MIC anti-biofilm activity is a property shared by very few antimicrobial agents.
Clinical Applications
Wound Healing Clinical Trials
The most advanced clinical application of LL-37 is in wound healing. The phase I/II trial by Gronberg et al. (2014) remains the only completed, published randomized controlled trial of LL-37 in humans. This first-in-man study enrolled 34 patients with hard-to-heal venous leg ulcers that had been present for at least 3 months despite standard care. Patients were randomized to receive either topical LL-37 (at doses of 0.5 mg/mL or 1.6 mg/mL) or placebo, applied directly to the wound bed twice weekly for 4 weeks.
The results were encouraging. LL-37-treated wounds showed significantly accelerated healing compared to placebo, with increased re-epithelialization rates and reduced wound area. No safety concerns were identified - neither local adverse events at the application site nor systemic adverse events were attributable to LL-37 treatment. These results provided the first clinical evidence supporting the safety and efficacy of exogenous LL-37 therapy in humans.
Cancer Immunotherapy Applications
LL-37 has entered early clinical testing for cancer applications. A phase I clinical trial evaluated intratumoral injection of LL-37 in melanoma patients with cutaneous metastases. The rationale was based on LL-37's ability to activate plasmacytoid dendritic cells, stimulating local anti-tumor immune responses. While full results have not been widely published, the trial demonstrated safety and provided proof-of-concept for LL-37 as a tumor immunostimulant.
Preclinical research has explored LL-37's anti-cancer properties in multiple tumor types. In pancreatic cancer, LL-37 inhibited tumor growth both in vitro and in vivo by inducing DNA damage, cell cycle arrest (G0/G1 phase), and reactive oxygen species production. In ovarian cancer, LL-37 enhanced the antitumor effects of CpG oligodeoxynucleotides, a TLR9 agonist used in cancer immunotherapy. Novel delivery systems, including LL-37-loaded zinc oxide nanoparticles and thermosensitive hydrogel nanoparticles, have shown improved antitumor activity in lung cancer models (Ren et al., 2022).
However, the relationship between LL-37 and cancer is not straightforward. In some cancer types, LL-37 appears to promote rather than inhibit tumor growth. In hepatocellular carcinoma, cathelicidin hCAP18/LL-37 promoted cell proliferation and suppressed the antitumor activity of vitamin D. In lung cancer, myeloid cell-derived LL-37 promoted tumor growth by activating Wnt/beta-catenin signaling. These conflicting findings highlight the context-dependent nature of LL-37's effects and caution against blanket therapeutic applications without consideration of tumor type and microenvironment.
Respiratory Infection Applications
The strong preclinical evidence for LL-37's antiviral activity, particularly against influenza and RSV, has generated interest in respiratory infection applications. The vitamin D-LL-37 axis provides a potential explanation for the well-documented association between vitamin D deficiency and respiratory infection susceptibility. Several clinical trials have tested vitamin D supplementation as an indirect strategy to boost LL-37 levels and reduce respiratory infections, with mixed but generally positive results in vitamin D-deficient populations.
A small-scale exploratory study evaluated oral LL-37 in COVID-19 patients, aiming to assess safety and preliminary efficacy. While the study was limited in size and design, it provided initial safety data for oral administration of LL-37 in the context of active viral infection. The rationale was based on LL-37's demonstrated antiviral activity against enveloped viruses and its immune-modulating properties that could potentially reduce the excessive inflammatory response seen in severe COVID-19.
Gut Health and Inflammatory Bowel Disease
LL-37's role in gastrointestinal health is an area of active investigation. The peptide is expressed by colonic epithelial cells, where it contributes to mucosal defense against enteric pathogens. Expression of LL-37 mRNA is significantly increased in the inflamed mucosa of both ulcerative colitis and Crohn's disease patients, suggesting a role in the mucosal immune response to intestinal inflammation (Schauber et al., 2006).
In mouse models of colitis, administration of LL-37 or its fragment KR-12 reduced intestinal inflammation and improved mucosal healing. The mechanisms appear to involve multiple pathways: direct antimicrobial activity against pathogenic gut bacteria, strengthening of the intestinal epithelial barrier, regulation of inflammatory cytokine production, and promotion of beneficial shifts in gut microbiota composition.
For those interested in gut health peptides, Larazotide targets the gut barrier through a different mechanism - regulation of tight junction permeability - and may complement LL-37's antimicrobial and anti-inflammatory gut activities. The anti-inflammatory peptide KPV also shows significant gut-directed anti-inflammatory properties.
Dermatological Applications
The skin is both a major production site and a key therapeutic target for LL-37. In healthy skin, constitutive LL-37 expression provides baseline antimicrobial defense. In atopic dermatitis (eczema), LL-37 expression is characteristically reduced, which may contribute to the increased susceptibility to skin infections (particularly S. aureus colonization) seen in these patients. Conversely, in rosacea, LL-37 is aberrantly processed into pro-inflammatory forms that contribute to the disease's characteristic facial redness and inflammation (Yamasaki et al., 2007).
These observations have led to interest in both boosting LL-37 (for atopic dermatitis and wound healing) and modulating its processing (for rosacea) as dermatological therapeutic strategies. The vitamin D analogue calcipotriol, already used topically for psoriasis, has been shown to enhance hCAP18/LL-37 upregulation during wounding in human skin, potentially offering a dual benefit of vitamin D receptor activation and enhanced antimicrobial defense.

Dosing Protocols and Administration
Research Dosing Guidelines
It is essential to emphasize that LL-37 dosing information presented here is based on published research protocols and community reports. There are no FDA-approved indications for exogenous LL-37 administration, and all dosing should be supervised by a qualified healthcare provider. Individuals seeking personalized guidance can take the Free Assessment at FormBlends to help determine appropriate peptide protocols.
The existing dosing literature for subcutaneous LL-37 administration can be summarized across several tiers:
| Protocol Level | Dose per Injection | Frequency | Cycle Length | Notes |
|---|---|---|---|---|
| Conservative/Beginner | 100 microg | Once daily, 5 days/week | 2-4 weeks on, 2 weeks off | Assess tolerance before increasing |
| Standard Research Protocol | 100-200 microg | Once daily | 4-8 weeks on, 2-4 weeks off | Most commonly referenced protocol |
| Advanced/Acute Support | 200-400 microg | Once daily | 2-4 weeks | Used during active immune challenges |
| Topical (Wound Healing) | 0.5-1.6 mg/mL | Twice weekly | 4 weeks | Per Gronberg et al. clinical trial |
The Dosing Calculator at FormBlends can help calculate reconstitution volumes and injection volumes based on vial concentration and target dose. Standard LL-37 research vials typically contain 5 mg of lyophilized peptide that requires reconstitution with bacteriostatic water before use.
Reconstitution and Storage
LL-37 is supplied as a lyophilized (freeze-dried) powder that must be reconstituted before use. The standard reconstitution procedure involves:
- Allow the vial to reach room temperature before opening
- Add bacteriostatic water slowly down the side of the vial (not directly onto the powder)
- Gently swirl to dissolve - do not shake vigorously, as this can denature the peptide
- Once reconstituted, store at 2-8 degrees Celsius (standard refrigerator temperature)
- Use within 4-6 weeks of reconstitution
For a 5 mg vial reconstituted with 2 mL of bacteriostatic water, each 0.1 mL (10 units on an insulin syringe) delivers 250 micrograms. For lower doses, reconstituting with more water allows more precise dosing. For example, adding 5 mL to a 5 mg vial creates a 1 mg/mL solution where each 0.1 mL delivers 100 micrograms.
Timing and Administration Considerations
Subcutaneous injection is the most common administration route for research applications. Injection sites include the abdominal area (rotating injection sites), the deltoid region, or the thigh. Injection-site reactions including mild burning, redness, or swelling are the most commonly reported side effects and are generally mild and self-limiting.
Some research protocols recommend morning administration, as LL-37 production follows a circadian rhythm that peaks during daytime hours. However, there is no strong clinical evidence that timing significantly affects outcomes, and consistency of administration schedule is likely more valuable than specific timing.
For immune support applications, many research protocols suggest concurrent vitamin D supplementation to optimize endogenous LL-37 production alongside exogenous administration. A target 25(OH)D level of 40-60 ng/mL is commonly recommended in the research community to support maximal endogenous cathelicidin expression.
Safety Profile
Clinical Safety Data
The safety profile of LL-37 has been evaluated in both the wound healing clinical trial and various preclinical studies. The Gronberg et al. (2014) phase I/II trial in 34 venous leg ulcer patients reported no safety concerns regarding either local or systemic adverse events at topical doses of 0.5 mg/mL and 1.6 mg/mL applied twice weekly for 4 weeks. This represents the most comprehensive clinical safety data available for exogenous LL-37 administration.
Preclinical safety studies of synthetic LL-37 have shown excellent tolerability in in vitro immune assays and animal models. No hepatotoxicity, nephrotoxicity, or genotoxicity has been observed at therapeutically relevant doses. The peptide is rapidly degraded by endogenous proteases in plasma (half-life of approximately 15-30 minutes in circulation), which limits systemic exposure and reduces the risk of cumulative toxicity but also necessitates local or frequent administration to maintain therapeutic levels.
Reported Adverse Effects
The most commonly reported adverse effects from subcutaneous LL-37 administration include:
- Injection site reactions: Localized burning, redness, or swelling at the injection site. These are generally mild and resolve within hours. They are thought to result from LL-37's mast cell degranulating activity, which releases histamine locally.
- Flu-like symptoms: Occasionally reported at higher doses (above 300 micrograms per day), including mild fever, fatigue, and myalgia. These likely reflect LL-37's immunostimulatory effects and are typically self-limiting.
- Gastrointestinal effects: Rare reports of mild GI discomfort, particularly at higher doses, possibly related to LL-37's effects on gut microbiota or mast cell activation in the GI tract.
Contraindications and Precautions
While comprehensive contraindication data is limited due to the early stage of clinical development, several precautions are supported by the available research:
- Autoimmune conditions: Given LL-37's role in amplifying immune responses and its demonstrated involvement in psoriasis pathogenesis (via TLR9-mediated self-DNA recognition), individuals with active autoimmune conditions should exercise caution. LL-37 has been implicated in the pathophysiology of systemic lupus erythematosus, where it can form immune complexes with self-DNA that activate autoimmune responses (Kahlenberg and Kaplan, 2013).
- Active cancer: Due to LL-37's context-dependent effects on tumor growth (promoting in some cancers, inhibiting in others), use in individuals with active malignancies should be approached with extreme caution and only under oncological supervision.
- Pregnancy and lactation: No safety data is available for LL-37 use during pregnancy or lactation. Given the lack of data, use during these periods is not recommended.
- Rosacea: Aberrant LL-37 processing contributes to rosacea pathology, and exogenous LL-37 administration could theoretically worsen this condition.
Safety Advisory
LL-37 is an investigational peptide without FDA approval for any clinical indication. All use should be under the supervision of a qualified healthcare provider. The safety data presented here is derived from limited clinical trials and preclinical studies. Long-term safety of exogenous LL-37 administration has not been established. Individuals with autoimmune conditions, active cancers, or rosacea should discuss potential risks with their healthcare provider before considering LL-37.
Comparison with Other Immune Peptides
LL-37 vs. Thymosin Alpha-1
Thymosin Alpha-1 (TA1) and LL-37 both support immune function, but they operate through fundamentally different mechanisms and at different levels of the immune hierarchy. TA1 primarily enhances adaptive immunity by promoting T cell maturation, differentiation, and function. It activates dendritic cells, enhances antigen presentation, and supports the development of both Th1 and cytotoxic T cell responses. LL-37, by contrast, operates primarily at the innate immune level - directly killing pathogens, recruiting innate immune cells, and modulating the inflammatory response.
These complementary mechanisms make TA1 and LL-37 a logical combination for comprehensive immune support. LL-37 provides immediate, front-line defense through direct antimicrobial activity and innate immune cell recruitment, while TA1 builds and strengthens the adaptive immune responses needed for long-term pathogen clearance and immune memory. For detailed information on TA1's mechanisms and clinical evidence, see the Thymosin Alpha-1 research report.
LL-37 vs. BPC-157
BPC-157 shares LL-37's wound healing properties but works through largely distinct pathways. BPC-157 is a gastric pentadecapeptide that promotes tissue repair through nitric oxide system modulation, VEGF upregulation, and FAK-paxillin pathway activation. It is primarily studied for musculoskeletal injuries, GI tissue protection, and general tissue repair. LL-37's wound healing benefits are more closely tied to its antimicrobial activity (clearing wound infections that impede healing) and its direct effects on keratinocyte and fibroblast proliferation and migration.
In the context of wound healing, the combination addresses complementary challenges: LL-37 targets the infection/biofilm component while BPC-157 provides tissue-protective and pro-angiogenic support. The BPC-157 research report covers the extensive evidence base for this peptide.
LL-37 vs. KPV
KPV is a tripeptide derived from alpha-melanocyte-stimulating hormone (alpha-MSH) with potent anti-inflammatory properties. While both LL-37 and KPV modulate inflammation, they do so through different receptor systems and signaling pathways. KPV acts primarily through melanocortin receptors (MC1R) to suppress NF-kappaB-mediated inflammation. LL-37 modulates inflammation through FPRL-1, P2X7, and EGFR pathways, with the added benefit of direct antimicrobial activity.
For gut-specific applications, KPV's anti-inflammatory effects on colonocytes complement LL-37's antimicrobial and barrier-strengthening properties. Both peptides are being investigated for inflammatory bowel disease applications from different mechanistic angles.
LL-37 vs. TB-500
TB-500 (Thymosin Beta-4 fragment) and LL-37 both promote wound healing and tissue repair, but through very different molecular mechanisms. TB-500 works primarily through actin sequestration, promoting cell migration and reducing inflammation. LL-37 works through membrane disruption of pathogens, EGFR transactivation, and immune cell recruitment. For comprehensive TB-500 information, see the TB-500 research report.
| Feature | LL-37 | Thymosin Alpha-1 | BPC-157 | KPV | TB-500 |
|---|---|---|---|---|---|
| Primary Function | Antimicrobial defense | Adaptive immune enhancement | Tissue repair/protection | Anti-inflammation | Tissue repair/migration |
| Direct Antimicrobial | Yes (broad-spectrum) | Indirect | No | Some | No |
| Wound Healing | Strong | Moderate (indirect) | Very Strong | Moderate | Very Strong |
| Immune Modulation | Innate focus | Adaptive focus | Minimal | Anti-inflammatory | Moderate |
| Anti-Biofilm | Strong | No | No | No | No |
| Gut Health | Barrier + antimicrobial | Indirect | Strong (gastroprotective) | Strong (anti-inflammatory) | Moderate |
| Administration Route | SC injection, topical | SC injection | SC injection, oral | SC injection, oral, topical | SC injection |
LL-37 vs. GHK-Cu
GHK-Cu (copper peptide) promotes wound healing through copper-dependent mechanisms that support collagen synthesis, angiogenesis, and anti-inflammatory signaling. While both LL-37 and GHK-Cu promote wound healing, they target different aspects of the healing process. LL-37 addresses the infection and immune components, while GHK-Cu focuses on extracellular matrix remodeling and collagen deposition. In combination, they could address both the antimicrobial and structural repair aspects of wound healing simultaneously.
Stacking Protocols
Immune Defense Stack
For comprehensive immune support, a commonly discussed research protocol combines LL-37 with Thymosin Alpha-1. This combination targets both arms of the immune system: LL-37 bolsters innate defenses through direct antimicrobial action and innate immune cell modulation, while TA1 enhances adaptive immunity through T cell and dendritic cell support. This dual-arm approach provides more comprehensive immune coverage than either peptide alone.
A typical immune defense research protocol might include:
- LL-37: 100-200 microg subcutaneously, once daily, 5 days per week
- Thymosin Alpha-1: 1.6 mg subcutaneously, twice weekly
- Vitamin D3: 5,000-10,000 IU daily (to support endogenous LL-37 production)
- Cycle: 4-8 weeks on, 2-4 weeks off
Wound Healing and Tissue Repair Stack
For wound healing applications, LL-37 pairs logically with BPC-157 and/or TB-500. LL-37 addresses the antimicrobial and immune aspects of wound healing (clearing infections, disrupting biofilms, recruiting immune cells), while BPC-157 and TB-500 promote tissue repair through angiogenesis, cell migration, and extracellular matrix remodeling.
A research-based wound healing protocol might include:
- LL-37: 100-200 microg subcutaneously, once daily
- BPC-157: 250-500 microg subcutaneously, once or twice daily
- TB-500: 2-5 mg subcutaneously, twice weekly
- Cycle: 4-6 weeks, reassess based on healing progress
Gut Health and Barrier Repair Stack
For gut-specific applications, LL-37 combines well with KPV and Larazotide. LL-37 provides antimicrobial defense and immune modulation at the gut mucosal surface, KPV delivers potent anti-inflammatory signaling to reduce intestinal inflammation, and Larazotide targets tight junction regulation to restore barrier integrity.
A gut-focused research protocol might include:
- LL-37: 100-200 microg subcutaneously, once daily
- KPV: 200-500 microg subcutaneously or orally, once daily
- Larazotide: per published protocol (typically oral administration)
- BPC-157: 250-500 microg orally, twice daily (for gastroprotection)
- Cycle: 4-8 weeks, monitor symptoms
Skin Health and Dermatology Stack
For dermatological applications, LL-37 combines with GHK-Cu for comprehensive skin health support. LL-37 provides antimicrobial defense and promotes keratinocyte migration, while GHK-Cu supports collagen synthesis and skin remodeling.
Stacking Disclaimer
All stacking protocols described here are based on research literature and community reports. They are not FDA-approved treatment regimens. Combining multiple peptides increases both the potential benefits and the complexity of monitoring for adverse effects. Any multi-peptide protocol should be supervised by a knowledgeable healthcare provider. Visit FormBlends Free Assessment for personalized guidance.
LL-37 and Neuroimmunomodulation
The Brain-Immune Connection
The connection between LL-37 and the nervous system represents an emerging area of research. LL-37 is expressed in the central nervous system, particularly in the brain's choroid plexus and in microglia (the brain's resident immune cells). Its expression in neural tissue suggests a role in neuroimmune defense against potential CNS infections, and recent work has begun to explore whether LL-37 may have neuromodulatory properties beyond pure antimicrobial function.
For researchers interested in the intersection of immune and neurological peptide effects, Selank represents a complementary approach. Selank is a synthetic peptide analog of tuftsin (a naturally occurring immunomodulatory peptide) that has been shown to have both anxiolytic and immunomodulatory properties. While LL-37 operates primarily at the innate immune level with secondary neurological effects, Selank works primarily through neurological pathways with secondary immune-modulating properties.
Future Research Directions
Next-Generation LL-37 Analogs
One of the major challenges limiting LL-37's clinical application is its relatively low proteolytic stability. The peptide is rapidly degraded by serum proteases, with a circulating half-life of approximately 15-30 minutes. This necessitates either frequent dosing or local application to maintain therapeutic concentrations. Researchers are actively developing modified versions of LL-37 designed to overcome this limitation.
Several approaches are being explored. D-amino acid substitution - replacing specific L-amino acids with their mirror-image D-forms - dramatically increases protease resistance while often maintaining antimicrobial activity. Cyclization of the peptide backbone can also improve stability. Truncated analogs like IG-25 (residues 13-37), GF-17 (residues 17-32), and KR-12 (residues 18-29) have been identified as minimum active fragments that retain antimicrobial activity with potentially improved therapeutic properties.
The LL-37 analog P60.4Ac has shown particular promise. This modified peptide retains broad-spectrum antimicrobial and anti-biofilm activity while demonstrating improved stability and reduced cytotoxicity compared to the parent molecule. P60.4Ac and the related peptide P10 have been shown to eradicate methicillin-resistant S. aureus from thermally wounded human skin equivalents, supporting their development for burn wound applications (de Breij et al., 2015).
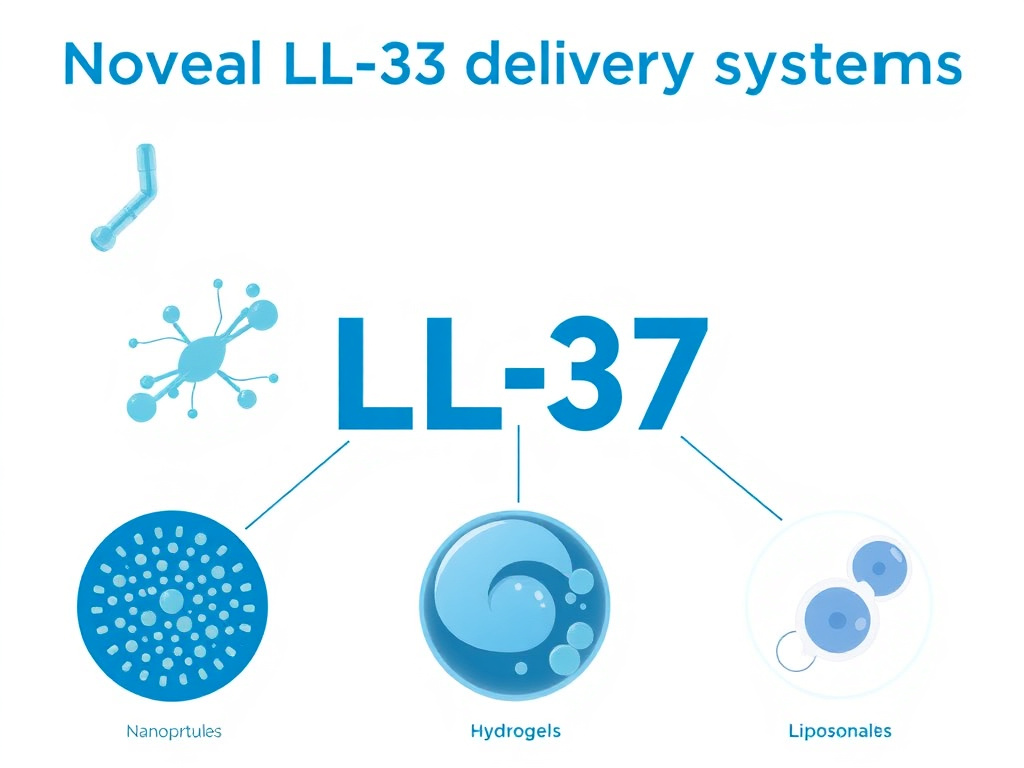
Novel Delivery Systems
Advanced delivery technologies are being developed to address LL-37's stability limitations and enable targeted delivery to specific tissues. These include:
- Hydrogel formulations: LL-37-loaded thermosensitive hydrogels that gel at body temperature, providing sustained peptide release at wound sites
- Nanoparticle carriers: LL-37 conjugated to zinc oxide or iron oxide nanoparticles for combined antimicrobial and nanoparticle-mediated effects
- Liposomal encapsulation: LL-37 packaged in liposomes for protected delivery and enhanced cellular uptake
- Polymer conjugation: PEGylation and other polymer conjugation strategies to extend circulating half-life
- Gene therapy approaches: Delivery of the CAMP gene to target tissues for sustained endogenous LL-37 production
Combination Therapy with Conventional Antibiotics
A particularly promising research direction involves combining LL-37 or its analogs with conventional antibiotics. The rationale is compelling: LL-37's membrane-disrupting activity can increase bacterial permeability to antibiotics that normally cannot cross the cell membrane efficiently. Several studies have demonstrated cooperative interactions between LL-37 and antibiotics including vancomycin, rifampin, ciprofloxacin, and polymyxin B. This approach could potentially rescue antibiotics that have lost effectiveness due to resistance, reduce the doses of antibiotics needed (reducing side effects), and extend the clinical useful life of existing antibiotic classes.
Frequently Asked Questions
LL-37 Against Gram-Negative Pathogens: Detailed Mechanisms
LPS Neutralization and Endotoxin Binding
One of LL-37's most clinically valuable properties against Gram-negative bacteria extends beyond simple killing - the peptide directly neutralizes bacterial lipopolysaccharide (LPS), also known as endotoxin. LPS is one of the most potent activators of the innate immune system. When released from dying Gram-negative bacteria during infection or antibiotic treatment, free LPS can trigger a catastrophic inflammatory cascade that leads to septic shock, multi-organ failure, and death. LL-37 binds LPS with high affinity, preventing it from activating TLR4/MD-2 signaling complexes on immune cells.
The LPS-binding capacity of LL-37 has been quantified in multiple studies. At concentrations achievable in vivo during active infection, LL-37 can neutralize LPS-induced TNF-alpha production from macrophages by 50-80%. This dual activity - killing the bacteria while simultaneously neutralizing the endotoxin they release - makes LL-37 conceptually superior to antibiotics for Gram-negative sepsis, since antibiotics kill bacteria but can actually worsen endotoxemia by releasing large amounts of LPS from lysed bacterial cells.
The mechanism of LPS binding involves electrostatic interactions between LL-37's positively charged residues and the negatively charged phosphate groups of lipid A, the toxic moiety of LPS. Once bound, the LL-37-LPS complex adopts a conformation that prevents LPS from interacting with LPS-binding protein (LBP) and CD14, the upstream components of the TLR4 signaling pathway. This means that even if LL-37 concentrations are insufficient for direct bacterial killing, the peptide can still provide significant protection against endotoxin-mediated immune activation.
Animal studies have provided compelling evidence for LL-37's anti-sepsis potential. In murine sepsis models, administration of LL-37 or the murine cathelicidin mCRAMP significantly improved survival rates, reduced circulating endotoxin levels, and attenuated organ damage compared to untreated controls. These findings, combined with LL-37's broad-spectrum antibacterial activity, have positioned cathelicidin-based therapies as potential adjuncts to conventional antibiotic treatment in sepsis management (Nagaoka et al., 2020).
Specific Interactions with Pseudomonas aeruginosa
P. aeruginosa deserves special attention because it is one of the most clinically significant Gram-negative pathogens and one that has developed extensive resistance to conventional antibiotics. This organism causes severe infections in immunocompromised patients, burn victims, and individuals with cystic fibrosis (CF). In CF patients, chronic P. aeruginosa lung infections are a leading cause of morbidity and mortality.
LL-37's interactions with P. aeruginosa are complex and multi-layered. At bactericidal concentrations (MIC typically 4-16 micrograms/mL), the peptide directly kills planktonic P. aeruginosa through membrane disruption. But at sub-MIC concentrations, LL-37 exerts at least four distinct anti-virulence effects: it inhibits biofilm formation, reduces swimming and swarming motility, downregulates quorum sensing gene expression, and decreases the production of elastase and other virulence factors.
The CF lung presents a particularly challenging environment for LL-37 activity. The high salt concentration in CF airway surface liquid (due to dysfunctional CFTR chloride channels) partially inhibits LL-37's antimicrobial activity, as elevated NaCl concentrations interfere with the electrostatic interactions that drive membrane binding. Additionally, P. aeruginosa strains isolated from chronic CF infections often show increased resistance to LL-37 compared to non-CF isolates, suggesting that chronic exposure can select for partial resistance mechanisms including LPS modifications that reduce negative surface charge.
Despite these challenges, LL-37 analogs designed for improved salt tolerance represent a promising therapeutic strategy for CF-related infections. Several modified peptides based on the LL-37 scaffold have been developed with enhanced antimicrobial activity in high-salt conditions, potentially overcoming one of the major limitations of native LL-37 in the CF lung environment.
Resistance Mechanisms in Gram-Negative Bacteria
While bacteria have great difficulty developing full resistance to LL-37's multi-target mechanism of action, certain Gram-negative species have evolved partial defense strategies. Understanding these mechanisms is valuable for predicting therapeutic limitations and designing improved analogs.
LPS modification is the most common strategy. Several bacterial species can alter the charge or structure of their LPS to reduce LL-37 binding. Salmonella typhimurium, for example, uses the PhoP/PhoQ two-component regulatory system to modify lipid A by adding aminoarabinose residues, which partially neutralize the negative charge and reduce LL-37's electrostatic attraction. Similar modifications have been identified in P. aeruginosa, Klebsiella pneumoniae, and other species.
Protease production is another defense strategy. Some Gram-negative bacteria produce outer membrane proteases capable of degrading LL-37 before it reaches lethal concentrations at the cell surface. P. aeruginosa's elastase and alkaline protease can both cleave LL-37, though the peptide must be present at relatively low concentrations for this defense to be effective - at higher concentrations, the antimicrobial activity overwhelms the proteolytic capacity.
Efflux pumps, which are well-known mediators of antibiotic resistance, play a limited role in LL-37 resistance. While some studies have shown that efflux pump overexpression can modestly reduce susceptibility to LL-37, the effect is small compared to the profound resistance these pumps confer against small-molecule antibiotics. This is likely because LL-37's primary target is the outer membrane itself, which the peptide encounters before reaching the efflux pump substrates.
These partial resistance mechanisms rarely confer more than a 2-4 fold increase in MIC, in contrast to the 100-1000 fold increases commonly seen with antibiotic resistance. This difference reflects the fundamental challenge bacteria face in resisting membrane-active peptides: they cannot fundamentally redesign their membrane architecture without compromising essential cellular functions.
LL-37 Against Gram-Positive Pathogens
Staphylococcal Infections and LL-37
Staphylococcus aureus is one of the most important human pathogens, causing infections ranging from minor skin abscesses to life-threatening bacteremia, endocarditis, and necrotizing pneumonia. The rise of methicillin-resistant S. aureus (MRSA) has created an urgent need for alternative antimicrobial approaches, and LL-37's retained activity against MRSA makes it a compelling candidate.
The mechanism of LL-37's action against S. aureus involves initial binding to the cell wall teichoic acids and lipoteichoic acids, which serve as the Gram-positive equivalent of LPS as negatively charged surface components. After traversing the thick peptidoglycan cell wall, LL-37 reaches the cytoplasmic membrane, where it exerts its membrane-disrupting effects. The thick cell wall of Gram-positive bacteria provides some protection by slowing peptide transit, which partially explains why MIC values against S. aureus (2-8 micrograms/mL) are generally slightly higher than those against E. coli (1-4 micrograms/mL).
Research on LL-37's effects against S. aureus virulence factors has revealed additional therapeutic benefits beyond direct killing. Sub-MIC concentrations of LL-37 reduce the production of several S. aureus toxins including alpha-hemolysin, Panton-Valentine leukocidin (PVL), and toxic shock syndrome toxin-1 (TSST-1). These toxins are major contributors to the severity of staphylococcal infections, and their downregulation by LL-37 could attenuate disease severity even in cases where complete bacterial eradication is not achieved.
In the context of skin and soft tissue infections, the most common manifestation of S. aureus disease, LL-37 provides a multi-pronged defense. It kills planktonic bacteria, disrupts surface biofilms, reduces toxin production, recruits neutrophils and other immune cells to the infection site, and promotes wound healing in the damaged tissue. This combination of activities makes LL-37 a natural defense molecule exquisitely designed for combating superficial staphylococcal infections.
Streptococcal Defense
Group A Streptococcus (Streptococcus pyogenes) causes a wide spectrum of diseases from pharyngitis ("strep throat") and impetigo to severe invasive infections like necrotizing fasciitis and streptococcal toxic shock syndrome. LL-37 shows strong activity against S. pyogenes with MIC values typically in the range of 1-4 micrograms/mL, making it one of the most susceptible Gram-positive pathogens.
However, S. pyogenes has evolved several countermeasures against LL-37. The bacterium produces the protease SpeB (streptococcal pyrogenic exotoxin B), which can cleave and inactivate LL-37. It also produces the M protein, a surface fibrillar protein that can bind and sequester LL-37, reducing its effective concentration at the bacterial surface. Some strains express the protease ScpC, which cleaves LL-37 at specific sites. These resistance mechanisms highlight the ongoing evolutionary arms race between human antimicrobial peptides and bacterial pathogens.
Despite these countermeasures, LL-37 remains an effective anti-streptococcal agent in most clinical contexts. The concentrations achieved at mucosal surfaces during active infection typically exceed the levels at which protease-mediated degradation can significantly impact LL-37 activity. And in individuals with adequate vitamin D levels, the continuous production of LL-37 provides a sustained antimicrobial pressure that helps prevent streptococcal colonization from progressing to symptomatic infection.
The Enterococcal Challenge
Enterococci, particularly E. faecalis and E. faecium, represent a growing clinical challenge due to their intrinsic resistance to many antibiotics and their ability to acquire additional resistance mechanisms, including vancomycin resistance (VRE). LL-37's activity against enterococci is moderate, with MIC values typically in the range of 8-32 micrograms/mL, higher than those for staphylococci and streptococci.
The relatively lower potency against enterococci reflects several factors. Enterococcal cell membranes have a higher proportion of zwitterionic phospholipids compared to other Gram-positive bacteria, reducing the electrostatic attraction for cationic peptides. Some enterococcal species also produce the metalloprotease GelE, which can degrade LL-37. Additionally, enterococci can modify their membrane lipid composition in response to antimicrobial peptide exposure, further reducing susceptibility.
Despite these limitations, LL-37 retains activity against vancomycin-resistant enterococci at concentrations only modestly higher than those needed for vancomycin-susceptible strains. This is clinically significant because VRE infections are extremely difficult to treat with conventional antibiotics, and any agent that retains activity regardless of vancomycin resistance status has potential therapeutic value.
LL-37 in Dental and Oral Health
LL-37 in the Oral Cavity
The oral cavity is one of the most microbiologically complex environments in the human body, harboring over 700 bacterial species in various ecological niches. LL-37 plays a significant role in maintaining oral health by contributing to saliva's antimicrobial properties and defending against periodontal pathogens. The peptide is present in saliva, gingival crevicular fluid, and the junctional epithelium that connects teeth to surrounding tissue.
Salivary LL-37 concentrations in healthy adults typically range from 0.5 to 3 micrograms per milliliter. While these concentrations are below the MIC for many oral pathogens when tested in standard laboratory conditions, the oral environment differs significantly from laboratory assays. In the thin layer of saliva coating mucosal surfaces, effective LL-37 concentrations may be substantially higher than bulk salivary measurements suggest. Additionally, LL-37 works cooperatively with other salivary antimicrobial factors including lysozyme, lactoferrin, histatins, and defensins.
In periodontal disease, LL-37 expression is significantly upregulated in the gingival epithelium surrounding inflamed periodontal pockets. This increased expression represents the body's attempt to control the bacterial population in the subgingival space. However, the efficacy of this response is compromised by the proteolytic enzymes produced by periodontal pathogens, particularly Porphyromonas gingivalis, which produces potent proteases (gingipains) capable of degrading LL-37.
Periodontal Disease Applications
The relationship between LL-37 and periodontal disease is bidirectional and complex. On one hand, LL-37 provides antimicrobial defense against periodontal pathogens. On the other hand, certain periodontal bacteria have evolved mechanisms to evade or inactivate LL-37, and the peptide's pro-inflammatory properties may contribute to the tissue destruction characteristic of advanced periodontitis.
P. gingivalis, the keystone pathogen in periodontitis, has developed a particularly sophisticated relationship with LL-37. The bacterium's gingipain proteases (RgpA, RgpB, and Kgp) efficiently degrade LL-37, reducing its antimicrobial activity. At the same time, some LL-37 degradation products may retain partial antimicrobial or immunomodulatory activities, creating a complex interplay between pathogen and host defense.
Research into LL-37-based dental therapeutics has explored several approaches. Locally delivered LL-37 or its analogs in slow-release formulations could provide sustained antimicrobial protection in periodontal pockets. LL-37-coated dental implants have been studied as a strategy to prevent peri-implant infections (peri-implantitis), one of the most significant complications of dental implant therapy. And vitamin D supplementation to boost endogenous LL-37 production in the oral epithelium is being investigated as a preventive strategy for periodontal disease.
The anti-biofilm properties of LL-37 are particularly relevant in the oral context, where dental plaque is essentially a highly organized, multi-species biofilm. LL-37's ability to prevent initial bacterial attachment to surfaces, disrupt quorum sensing, and penetrate established biofilms could make it valuable for both preventing and treating dental plaque-related diseases.
LL-37 in Respiratory Health: A Detailed Analysis
LL-37 in Airway Surface Liquid
The respiratory tract is continuously exposed to inhaled pathogens, and the airway surface liquid (ASL) that lines the respiratory epithelium serves as the first line of defense. LL-37 is a key component of ASL antimicrobial activity, produced by airway epithelial cells and deposited by neutrophils that patrol the airway surface.
In healthy airways, LL-37 concentrations in ASL are estimated at 2-5 micrograms per milliliter, sufficient to provide baseline antimicrobial activity against common respiratory pathogens. During respiratory infections, local LL-37 levels can increase 5-10 fold through a combination of increased epithelial production (stimulated by inflammatory cytokines and TLR activation) and neutrophil influx and degranulation.
The respiratory epithelium's production of LL-37 is regulated by multiple factors. Vitamin D is the most well-characterized regulator, with calcitriol directly inducing CAMP gene transcription in bronchial and alveolar epithelial cells. Infection-related signals including IL-1beta, IL-17, and bacterial products acting through TLR2 and TLR5 also upregulate LL-37 production. This multi-input regulation ensures that LL-37 production scales with the level of microbial threat.
Tuberculosis and Mycobacterial Defense
The relationship between LL-37 and tuberculosis (TB) is one of the most compelling examples of the vitamin D-cathelicidin immune axis. Mycobacterium tuberculosis has a unique ability to survive inside macrophages, the very immune cells designed to kill intracellular pathogens. LL-37 plays a critical role in macrophage-mediated killing of M. tuberculosis, and this function is directly dependent on vitamin D status.
The discovery of this mechanism, reported by Liu et al. in 2006, was a landmark in immunology. The researchers found that TLR activation in macrophages upregulates the vitamin D receptor (VDR) and the enzyme CYP27B1, which converts circulating 25-hydroxyvitamin D to the active 1,25-dihydroxyvitamin D3 form. This locally produced active vitamin D then induces CAMP gene expression, leading to LL-37 production. The LL-37 is delivered to M. tuberculosis-containing phagosomes, where it contributes to mycobacterial killing.
This mechanism provided a molecular explanation for several longstanding clinical observations. The historical use of cod liver oil (rich in vitamin D) for TB treatment, the increased TB risk in vitamin D-deficient populations, and the seasonal variation in TB incidence all found a mechanistic basis in the vitamin D-cathelicidin-TB axis. Clinical trials of vitamin D supplementation in TB patients have shown enhanced sputum culture conversion rates in some studies, though results have been inconsistent across trials, likely reflecting differences in baseline vitamin D status and genetic variation in the VDR.
Influenza Defense: Detailed Mechanisms
LL-37's activity against influenza viruses has been studied more extensively than its activity against any other virus. The work of multiple research groups has revealed that LL-37 combats influenza through at least three distinct mechanisms.
First, direct virucidal activity: LL-37 binds to the influenza viral envelope and disrupts its integrity. Electron microscopy studies show viral particles treated with LL-37 display membrane blebbing, fragmentation, and loss of structural organization. This direct disruption is concentration-dependent and results in loss of viral infectivity.
Second, inhibition of viral attachment: LL-37 interferes with hemagglutinin-mediated binding of influenza virus to sialic acid residues on host cell surfaces. This prevents the initial attachment step that is required for viral entry. The mechanism appears to involve direct interaction between LL-37 and viral hemagglutinin, though the exact binding site has not been fully characterized.
Third, enhancement of host antiviral responses: LL-37 stimulates the production of type I interferons and interferon-stimulated genes in respiratory epithelial cells, enhancing the cellular antiviral state. This effect operates through LL-37's ability to deliver viral nucleic acids to endosomal pattern recognition receptors, amplifying the innate immune detection of viral infection.
The combined potency of these mechanisms was demonstrated dramatically in the mouse studies by Barlow et al. (2011), where LL-37 treatment reduced lung viral titers by 70-80% - comparable to zanamivir, a prescription neuraminidase inhibitor. Of note, LL-37's mechanism of action is entirely different from that of neuraminidase inhibitors, suggesting that combination therapy could potentially provide additive or cooperative antiviral effects.
The activity of LL-37 against pandemic influenza strains is particularly relevant. Tripathi et al. (2015) showed that LL-37 and derived peptides were effective against both seasonal and pandemic H1N1 influenza A virus strains, including the 2009 pandemic strain. The peptides neutralized both seasonal and pandemic strains in vitro and showed improved activity when used in combination with conventional antiviral approaches.
Respiratory Syncytial Virus (RSV)
RSV is the most common cause of lower respiratory tract infections in infants and young children, and a significant cause of respiratory illness in elderly and immunocompromised adults. LL-37 has shown antiviral activity against RSV in polarized airway epithelial cells, which more closely model the in vivo airway environment than standard cell culture systems.
Currie et al. (2016) demonstrated that LL-37 inhibited RSV replication through both direct virucidal activity and enhancement of epithelial cell antiviral defenses. The peptide stimulated production of interferon-beta and several interferon-stimulated genes in RSV-infected cells, suggesting that it amplifies the host's natural antiviral response in addition to directly neutralizing viral particles.
The clinical relevance of these findings is supported by epidemiological observations. Vitamin D deficiency in infancy is associated with increased risk of severe RSV bronchiolitis, and cord blood 25-hydroxyvitamin D levels inversely correlate with RSV infection severity in the first year of life. While these associations are not proof of a causal role for LL-37, they are consistent with the hypothesis that vitamin D-mediated cathelicidin production contributes to defense against RSV.
LL-37 and Skin Biology
Skin Barrier and Antimicrobial Defense
The skin is the body's largest organ and its primary physical barrier against environmental pathogens. LL-37 is a critical component of the skin's chemical defense system, working alongside other antimicrobial peptides (human beta-defensins, dermcidin, psoriasin) to create an antimicrobial shield on the skin surface.
Keratinocytes, the predominant cell type in the epidermis, constitutively express low levels of LL-37 that provide baseline antimicrobial protection. Upon injury or infection, keratinocyte LL-37 expression increases dramatically through multiple signaling pathways. TLR2 activation by Gram-positive bacterial products (lipoteichoic acid, peptidoglycan), TLR4 activation by Gram-negative LPS, and IL-17 signaling from resident T cells all independently upregulate CAMP gene expression in keratinocytes.
The vitamin D pathway is particularly active in skin. Keratinocytes possess the complete enzymatic machinery to convert vitamin D precursors (7-dehydrocholesterol) to the active 1,25-dihydroxyvitamin D3 form, meaning that skin cells can produce vitamin D locally in response to UV radiation and then immediately use it to upregulate LL-37 production. This creates a direct sun exposure-to-antimicrobial defense pathway that operates independently of circulating vitamin D levels.
Sweat glands also contribute LL-37 to the skin surface. The peptide is secreted in sweat and concentrates on the skin as perspiration evaporates, potentially reaching antimicrobial concentrations in areas of heavy sweating. This mechanism may partially explain the empirical observation that physical activity is associated with improved skin health and reduced incidence of skin infections.
Atopic Dermatitis and LL-37 Deficiency
Atopic dermatitis (AD, or eczema) affects approximately 10-20% of children and 1-3% of adults worldwide. One of the characteristic features of AD is increased susceptibility to skin infections, particularly S. aureus colonization, which affects up to 90% of AD patients compared to only 5-30% of healthy individuals. A key factor in this susceptibility is the deficient expression of antimicrobial peptides, including LL-37, in AD skin.
In healthy skin, wounding or infection triggers a strong increase in LL-37 expression. In AD skin, this induction is markedly blunted. The mechanism involves the Th2 cytokine environment characteristic of AD - IL-4 and IL-13, the dominant cytokines in AD inflammation, actively suppress CAMP gene expression in keratinocytes. This creates a paradoxical situation where the very inflammatory process that should be mounting an antimicrobial defense is instead suppressing one of the skin's key antimicrobial effectors.
This understanding has therapeutic implications. Strategies to boost LL-37 levels in AD skin could potentially reduce S. aureus colonization and break the infection-inflammation cycle that drives disease flares. Vitamin D supplementation has been studied in AD patients with the specific goal of increasing cathelicidin levels, and several small studies have shown both increased LL-37 expression and clinical improvement with vitamin D treatment. Topical vitamin D analogs applied to AD-affected skin can also upregulate local LL-37 production.
Rosacea: When LL-37 Becomes Part of the Problem
Rosacea provides a fascinating counterpoint to atopic dermatitis - instead of too little LL-37, the disease involves abnormal processing of cathelicidin that generates pro-inflammatory peptide fragments. In rosacea-affected skin, the serine protease kallikrein 5 (KLK5) is overexpressed and aberrantly active. Instead of properly processing hCAP18 to the native LL-37 peptide, the overactive KLK5 generates abnormal cathelicidin fragments that trigger inflammation, vasodilation, and angiogenesis - the hallmarks of rosacea.
Yamasaki et al. (2007) demonstrated that injection of the abnormal cathelicidin fragments found in rosacea skin into mouse skin reproduced the clinical features of rosacea, while injection of native LL-37 did not cause the same inflammatory response. This work established cathelicidin processing abnormalities as a key pathogenic mechanism in rosacea and explained why the disease is associated with skin that appears "overreactive" to normally tolerated stimuli.
This finding has important implications for exogenous LL-37 use. Individuals with rosacea may need to exercise caution with LL-37 supplementation, as the additional cathelicidin could be aberrantly processed by the overactive KLK5 in their skin, potentially worsening symptoms. Conversely, strategies that normalize KLK5 activity rather than increasing total cathelicidin levels might be more appropriate for rosacea patients.
Psoriasis and the LL-37-DNA Complex
Psoriasis involves another pathological manifestation of LL-37 activity. In this autoimmune skin condition, dying keratinocytes release self-DNA that forms complexes with LL-37. These LL-37/self-DNA complexes are internalized by plasmacytoid dendritic cells and delivered to endosomal TLR9, which normally detects microbial DNA. The TLR9 activation triggers type I interferon production, which drives the characteristic psoriatic inflammatory cascade.
This mechanism, discovered by Lande et al. (2007), revealed that LL-37 can essentially break immune tolerance to self-DNA by serving as a delivery vehicle that converts harmless extracellular self-DNA into a potent TLR9 agonist. The same delivery mechanism that enhances antimicrobial immunity (by helping immune cells detect microbial DNA) becomes pathogenic when it delivers self-DNA instead.
The psoriasis connection has broader implications for understanding autoimmune disease. Similar LL-37/self-nucleic acid complexes have been implicated in the pathogenesis of systemic lupus erythematosus (SLE), where LL-37 forms complexes with self-DNA and self-RNA that activate both TLR9 and TLR7/8, driving the interferon signature characteristic of SLE. These findings underscore the dual-edged nature of LL-37's immunomodulatory properties - protective in the context of infection but potentially harmful in the context of tissue damage and self-antigen release.
LL-37 and the Gut Microbiome
Microbiome Shaping Effects
The gut microbiome is a complex ecosystem of trillions of microorganisms that plays essential roles in digestion, immune development, vitamin synthesis, and protection against pathogenic colonization. LL-37, produced by intestinal epithelial cells and Paneth cells in the small intestinal crypts, helps shape and maintain this microbial community through selective antimicrobial pressure.
Not all bacteria are equally susceptible to LL-37, and this differential susceptibility has microbiome-shaping implications. Many commensal (beneficial) bacteria have evolved mechanisms to tolerate LL-37 at concentrations that are lethal to many pathogenic species. Lactobacillus species, for example, show relatively high resistance to LL-37, while many enteric pathogens (pathogenic E. coli, Salmonella, Shigella) are more susceptible. This selective pressure helps maintain a healthy balance between commensal and pathogenic organisms in the gut.
The relationship between LL-37 and the gut microbiome is bidirectional. Short-chain fatty acids produced by gut bacteria, particularly butyrate from fiber fermentation, can induce CAMP gene expression in colonic epithelial cells. This means that a healthy, fiber-rich diet that promotes butyrate-producing bacteria also enhances local LL-37 production, creating a positive feedback loop between dietary fiber, beneficial gut bacteria, and antimicrobial defense.
Intestinal Barrier Integrity
Beyond its antimicrobial effects, LL-37 contributes to intestinal barrier integrity through direct effects on epithelial cell biology. The peptide promotes the proliferation and migration of intestinal epithelial cells, supporting the rapid turnover of the intestinal lining (the gut epithelium is completely replaced every 3-5 days). It also modulates tight junction protein expression, helping maintain the paracellular barrier that prevents uncontrolled passage of luminal contents into the underlying tissue.
In inflammatory bowel disease (IBD), both the antimicrobial and barrier-maintaining functions of LL-37 are relevant. Crohn's disease and ulcerative colitis are both characterized by disrupted barrier function, altered microbiome composition, and inappropriate immune responses to gut bacteria. LL-37 expression is increased in inflamed IBD mucosa, representing an attempt by the tissue to restore antimicrobial defense and barrier function. However, as with psoriasis, the pro-inflammatory aspects of LL-37 may contribute to perpetuating the inflammatory cycle in some contexts.
For individuals interested in gut barrier support, Larazotide targets tight junction regulation through a mechanism distinct from LL-37, while BPC-157 promotes gut mucosal healing through nitric oxide system modulation. The combination of LL-37 (antimicrobial defense), Larazotide (barrier regulation), BPC-157 (tissue repair), and KPV (anti-inflammation) represents a multi-targeted approach to gut health that addresses different facets of intestinal pathology.
Helicobacter pylori and Gastric Defense
H. pylori infects approximately half the world's population and causes gastritis, peptic ulcers, and gastric cancer. LL-37 is expressed in the gastric mucosa and shows direct antimicrobial activity against H. pylori with MIC values in the range of 2-8 micrograms/mL. The peptide is found in gastric juice and contributes to the stomach's antimicrobial defense alongside acid, pepsin, and other protective factors.
However, H. pylori has evolved several strategies to resist LL-37. The bacterium modifies its LPS to reduce LL-37 binding, produces proteases that can degrade the peptide, and creates a microenvironment within the gastric mucus layer that partially shields it from luminal antimicrobial factors. The chronic nature of H. pylori infection suggests that these evasion strategies are at least partially effective against endogenous LL-37 defense.
Research into LL-37-based therapeutics for H. pylori eradication is at an early stage but shows promise, particularly for antibiotic-resistant strains that are increasingly common. The combination of LL-37's direct anti-H. pylori activity with its wound healing and anti-inflammatory properties could make it valuable as an adjunct to standard triple or quadruple therapy regimens for H. pylori eradication.
Exercise, Lifestyle Factors, and LL-37 Levels
Exercise and Cathelicidin Production
Physical exercise influences LL-37 levels through multiple pathways. Moderate exercise increases circulating neutrophil counts (and therefore the total body reservoir of stored hCAP18/LL-37), improves vitamin D metabolism, and may directly upregulate CAMP gene expression in circulating immune cells. Studies have shown that moderate-intensity exercise leads to transient increases in plasma LL-37 concentrations, with peak levels occurring 1-2 hours post-exercise.
The relationship between exercise intensity and LL-37 levels follows an inverted U-curve. Moderate exercise appears to boost antimicrobial peptide production, while prolonged, exhaustive exercise (such as marathon running or intense training sessions lasting more than 90 minutes) can temporarily suppress LL-37 levels, contributing to the well-documented "open window" of increased infection susceptibility that follows intense physical exertion. This temporary immunosuppression typically resolves within 24-72 hours.
For athletes and physically active individuals, the exercise-LL-37 connection has practical implications. Maintaining adequate vitamin D status may help mitigate the post-exercise dip in antimicrobial defense by ensuring maximal CAMP gene responsiveness to vitamin D signaling. Avoiding vitamin D deficiency is particularly important for athletes who train indoors (limiting sun exposure) or who compete in northern latitudes during winter months.
Sleep, Stress, and Circadian Factors
LL-37 production follows a circadian rhythm, with higher levels during daytime hours and lower levels at night. This pattern aligns with the broader circadian regulation of immune function and may contribute to the time-of-day variation in infection susceptibility that has been observed in clinical studies.
Chronic stress and sleep deprivation both negatively impact LL-37 levels. Cortisol, the primary stress hormone, suppresses CAMP gene expression through glucocorticoid receptor-mediated transcriptional repression. Chronic sleep restriction (less than 6 hours per night) is associated with reduced circulating LL-37 levels and increased susceptibility to respiratory infections. These observations provide a molecular mechanism linking psychosocial stress and sleep quality to infection risk.
The practical takeaway is straightforward: lifestyle factors that support overall immune health also support LL-37 production. Adequate sleep (7-9 hours per night), stress management, regular moderate exercise, sun exposure or vitamin D supplementation, and a fiber-rich diet all contribute to maintaining optimal endogenous LL-37 levels. These lifestyle factors should be considered foundational to any peptide supplementation strategy, as they optimize the body's own antimicrobial peptide production.
Dietary Influences on LL-37
Beyond vitamin D, several dietary factors influence LL-37 production. Butyrate, a short-chain fatty acid produced by gut bacterial fermentation of dietary fiber, directly induces CAMP gene expression in colonic epithelial cells through histone deacetylase (HDAC) inhibition. This means that a high-fiber diet rich in resistant starch, inulin, and other fermentable fibers promotes butyrate production by commensal bacteria, which in turn enhances local LL-37 production in the gut epithelium.
Bile acids, produced by the liver from cholesterol and modified by gut bacteria, have also been shown to influence cathelicidin expression. Lithocholic acid (LCA), a secondary bile acid produced by gut bacterial metabolism, activates the vitamin D receptor and can induce CAMP gene expression independently of vitamin D. This provides yet another link between gut microbial metabolism and antimicrobial peptide production.
Zinc, an essential trace mineral, is required for optimal antimicrobial peptide production and function. Zinc deficiency is associated with reduced LL-37 levels and impaired antimicrobial defense. Zinc supplementation in deficient individuals can restore LL-37 production. Foods rich in zinc include oysters, red meat, poultry, beans, nuts, and whole grains.
The omega-3 fatty acids EPA and DHA, found in fatty fish and fish oil supplements, have complex effects on LL-37. Some studies show that omega-3 supplementation increases LL-37 expression in immune cells, while others suggest that the anti-inflammatory effects of omega-3s may indirectly reduce LL-37 production by dampening the inflammatory signals that upregulate CAMP gene expression. The net effect likely depends on the individual's baseline inflammatory status and dietary omega-6 to omega-3 ratio.

Drug Interactions and Practical Considerations
Antibiotic Cooperation and Combination Potential
The interaction between LL-37 and conventional antibiotics is an area of significant clinical interest. Multiple studies have demonstrated cooperative or additive interactions between LL-37 and several antibiotic classes, suggesting that LL-37 could potentially enhance the effectiveness of conventional antibiotic therapy.
The mechanisms underlying antibiotic cooperation vary depending on the antibiotic class. For antibiotics that must cross the bacterial membrane to reach their intracellular targets (aminoglycosides, fluoroquinolones, tetracyclines), LL-37's membrane-permeabilizing activity can increase intracellular antibiotic accumulation. This is particularly valuable for antibiotics whose efficacy is limited by poor membrane penetration or efflux pump-mediated removal.
For cell wall-active antibiotics (beta-lactams, vancomycin), the cooperative mechanism is different. LL-37 and cell wall-active antibiotics attack complementary targets - the cell wall and the cell membrane, respectively - creating a coordinated assault that bacteria find difficult to resist. Studies have shown that sub-MIC concentrations of LL-37 can reduce the MIC of methicillin for MRSA strains, potentially restoring susceptibility to a widely available and well-tolerated antibiotic class.
Polymyxin B, a cationic antimicrobial peptide antibiotic used as a last-resort treatment for multidrug-resistant Gram-negative infections, shows particularly strong cooperative effects with LL-37. The combination achieves biofilm eradication at concentrations where neither agent alone is effective, and the dual peptide approach appears to overwhelm the limited resistance mechanisms bacteria have evolved against individual cationic peptides.
Interactions with Immunosuppressive Medications
Patients on immunosuppressive medications require careful consideration before using LL-37. Corticosteroids, the most commonly used immunosuppressants, directly suppress CAMP gene expression through glucocorticoid receptor-mediated transcriptional repression. This means that patients on chronic corticosteroid therapy may have reduced endogenous LL-37 production, potentially benefiting from exogenous supplementation but also potentially experiencing altered responses to the peptide.
Calcineurin inhibitors (tacrolimus, cyclosporine), used in transplant medicine and for certain dermatological conditions, have complex effects on antimicrobial peptide expression. Some studies suggest that tacrolimus can partially restore cathelicidin expression in atopic dermatitis skin, while others show inhibitory effects on LL-37 production in other cell types. The net effect likely depends on the specific tissue and clinical context.
TNF-alpha inhibitors (infliximab, adalimumab, etanercept), widely used for autoimmune conditions, could theoretically interact with LL-37's immune-modulating effects. Since TNF-alpha is one of the cytokines that LL-37 can either promote or suppress depending on context, the addition of TNF-alpha blockade creates a complex immunological situation that has not been thoroughly studied. Caution is advised, and consultation with the prescribing physician is essential.
Practical Administration Timing
For individuals using LL-37 under medical supervision, several practical considerations can optimize outcomes. Morning administration aligns with the natural circadian peak of cathelicidin production and may provide the most physiologically congruent immune support. Injection approximately 30-60 minutes before or after exercise could theoretically complement the exercise-induced mobilization of immune cells, though this specific timing strategy has not been clinically validated.
Vitamin D supplementation is best taken with the largest meal of the day, as vitamin D is fat-soluble and its absorption is enhanced by dietary fat. If using LL-37 and vitamin D together, the goal is to achieve and maintain 25(OH)D levels in the 40-60 ng/mL range, which appears to optimize endogenous cathelicidin production without exceeding the saturation point identified by Dixon et al. (2012).
Rotation of injection sites is recommended to minimize injection site reactions, which are the most common side effect of subcutaneous LL-37 administration. The abdominal area offers the most surface area for rotation, and a systematic rotation pattern (upper left, upper right, lower left, lower right quadrants) can help ensure each site has adequate recovery time between injections.
Comparative Biology: Cathelicidins Across Species
Evolutionary Conservation of Cathelicidins
Cathelicidins are an ancient family of antimicrobial peptides found across virtually all vertebrate species. While humans produce only one cathelicidin (LL-37), many other species produce multiple cathelicidins with diverse structures and activities. Cattle produce over a dozen cathelicidins, pigs produce several (including the well-studied PR-39 and protegrin-1), and even fish and reptiles produce cathelicidin-like peptides. This widespread conservation across hundreds of millions of years of evolution underscores the fundamental importance of cathelicidins in host defense.
The murine cathelicidin CRAMP (cathelin-related antimicrobial peptide) is the closest mouse homolog to human LL-37. CRAMP shares approximately 67% sequence identity with LL-37 and displays similar antimicrobial and immunomodulatory properties. CRAMP-knockout mice show increased susceptibility to skin infections, wound infections, and experimental sepsis, confirming the essential role of cathelicidins in murine immunity and providing validated animal models for studying cathelicidin biology.
However, a critical difference exists between human and murine cathelicidin regulation. The vitamin D response element in the CAMP gene promoter is present only in primate genomes, not in mice or other rodents. This means that the vitamin D-cathelicidin axis - one of the most clinically significant aspects of LL-37 biology - cannot be studied in conventional mouse models. Researchers must use humanized mice, primate models, or in vitro human cell systems to investigate this regulatory pathway, which has limited the pace of translational research.
Veterinary Applications
The broad-spectrum antimicrobial properties of cathelicidins have generated interest in veterinary applications, particularly in the context of reducing antibiotic use in animal agriculture. Bovine cathelicidins (indolicidin, BMAP-27, BMAP-28) have been studied for treatment of bovine mastitis, one of the most common and economically significant infectious diseases in dairy cattle. Porcine cathelicidins have been investigated for controlling swine dysentery and other porcine enteric infections.
While these veterinary applications involve species-specific cathelicidins rather than human LL-37, the underlying principles are transferable. The mechanisms of membrane disruption, biofilm inhibition, and immune modulation are conserved across cathelicidin family members, and lessons learned from veterinary cathelicidin research inform the development of human LL-37-based therapeutics.
Quality Considerations and Sourcing
Peptide Purity and Authenticity
The quality of LL-37 preparations varies significantly between manufacturers, and this variation can substantially impact both efficacy and safety. Key quality parameters to consider include peptide purity (measured by HPLC), sequence accuracy (confirmed by mass spectrometry), endotoxin levels (measured by LAL assay), sterility (for injectable preparations), and proper storage and handling throughout the supply chain.
Research-grade LL-37 should have a purity of at least 95% as measured by reverse-phase HPLC. Lower-purity preparations may contain truncated peptides, oxidized variants, or synthesis byproducts that could have different biological activities or cause adverse reactions. Mass spectrometry confirmation of the correct molecular weight (4493.3 Da for LL-37) is essential to verify that the peptide has the correct amino acid sequence.
Endotoxin contamination is a particular concern for LL-37 preparations, as endotoxin from the manufacturing process can confound research results and cause febrile reactions in users. High-quality preparations should have endotoxin levels below 0.25 EU/mL, as measured by the Limulus Amebocyte Lysate (LAL) assay. FormBlends LL-37 provides third-party tested peptide with documented purity and quality specifications.
Storage and Stability
LL-37 stability is influenced by temperature, pH, light exposure, and the presence of oxidizing agents. The lyophilized (freeze-dried) form is the most stable and can be stored at -20 degrees Celsius for extended periods (typically 1-2 years) without significant degradation. At room temperature, lyophilized LL-37 may begin to degrade within weeks, particularly in humid environments.
Once reconstituted, LL-37 solutions are much less stable. At refrigerator temperatures (2-8 degrees Celsius), reconstituted LL-37 in bacteriostatic water typically maintains potency for 4-6 weeks. Repeated freeze-thaw cycles should be avoided, as they can cause peptide aggregation and loss of activity. For this reason, aliquoting reconstituted LL-37 into single-use portions can help maintain quality if the full vial won't be used within the recommended timeframe.
Light exposure, particularly UV light, can cause photo-oxidation of tryptophan and other susceptible amino acid residues in LL-37, reducing antimicrobial activity. Storing reconstituted LL-37 in amber vials or wrapping the vial in aluminum foil can protect against light-induced degradation.
Clinical Perspective and Summary
Current Evidence Landscape
The evidence base for LL-37 spans hundreds of in vitro studies, dozens of animal model investigations, and a small but growing number of human clinical trials. The quality and consistency of the preclinical evidence is strong: LL-37's antimicrobial, anti-biofilm, immunomodulatory, and wound healing properties have been replicated across multiple laboratories and model systems.
However, the clinical evidence remains limited. The only completed, published randomized controlled trial is the Gronberg et al. (2014) study of topical LL-37 for venous leg ulcers, which was a small phase I/II trial with 34 patients. While the results were encouraging, they represent preliminary evidence that requires confirmation in larger, longer-duration trials. The melanoma immunotherapy trial and the exploratory COVID-19 study provide additional safety data but limited efficacy evidence.
This evidence gap between strong preclinical data and limited clinical data is common for antimicrobial peptide therapeutics and reflects several challenges: the high cost of synthetic peptide production at pharmaceutical grade, the regulatory complexity of peptide drug development, the proteolytic instability of native LL-37, and the difficulty of designing clinical trials for a multi-functional agent that doesn't fit neatly into conventional drug categories.
Who Might Benefit Most from LL-37 Research
Based on the available evidence, the individuals most likely to benefit from LL-37 research applications include:
- Individuals with chronic or recurrent infections: Particularly those with biofilm-associated infections, recurrent skin and soft tissue infections, or chronic wound infections that have not responded to conventional antibiotic therapy
- Vitamin D-deficient individuals: Those with documented low 25(OH)D levels (below 30 ng/mL) who may have reduced endogenous LL-37 production, particularly if they also experience frequent infections
- People with chronic wounds: Including venous leg ulcers, diabetic foot ulcers, and pressure injuries, where the combination of antimicrobial and wound healing properties could provide combined benefits
- Immunocompromised individuals: Those with primary or secondary immunodeficiency who may benefit from enhanced innate immune defense (under appropriate medical supervision)
- People seeking comprehensive immune support: Particularly during cold and flu season or when facing increased infection risk due to travel, stress, or other factors
Practical Integration
For those considering LL-37 under medical supervision, a practical approach begins with optimizing endogenous LL-37 production through lifestyle and nutritional strategies: maintaining vitamin D levels in the 40-60 ng/mL range, consuming adequate dietary fiber to support butyrate production, getting sufficient sleep, managing stress, and engaging in regular moderate exercise. These foundational strategies support natural antimicrobial peptide production and should be implemented regardless of whether exogenous LL-37 supplementation is pursued.
For those who proceed to exogenous LL-37 use, starting with conservative doses (100 micrograms daily) and gradually increasing as tolerance is assessed provides the safest approach. Cycling (4-8 weeks on, 2-4 weeks off) is recommended based on the general principle of peptide cycling and the theoretical concern about prolonged immune stimulation. Monitoring for injection site reactions, flu-like symptoms, and any signs of autoimmune activation is advisable throughout the use period.
The Free Assessment at FormBlends provides personalized guidance on peptide selection and protocol design. For researchers and clinicians seeking current information on LL-37 and related compounds, the Science & Research section offers regularly updated resources.
Bottom Line
LL-37 stands at the intersection of antimicrobial defense, immune modulation, and tissue repair. As the sole human cathelicidin, it occupies a unique position in our immune system's architecture. The vitamin D connection provides a straightforward, evidence-based strategy for optimizing endogenous production. And while clinical development of exogenous LL-37 is still in early stages, the breadth and consistency of preclinical evidence - combined with favorable early clinical safety data - make this one of the most promising peptides in the antimicrobial and immune support research space. For those interested in exploring LL-37 further, FormBlends LL-37 offers research-grade peptide with documented quality specifications.
In-Depth: LL-37's Effects on Individual Immune Cell Populations
Neutrophil Modulation
Neutrophils occupy a unique position in the LL-37 story: they are both the largest producers and the most directly affected targets of this peptide. Each neutrophil stores approximately 630 ng of hCAP18 in its specific (secondary) granules, making neutrophil degranulation the fastest mechanism for releasing large quantities of LL-37 at sites of infection. But LL-37 also feeds back on neutrophils, modifying their behavior in ways that enhance antimicrobial function while limiting tissue damage.
At the functional level, LL-37 increases neutrophil phagocytic capacity. Neutrophils exposed to LL-37 engulf more bacteria per unit time compared to untreated cells, and they show enhanced oxidative burst activity (production of reactive oxygen species used for intracellular killing). Paradoxically, while enhancing these antimicrobial functions, LL-37 simultaneously reduces the release of pro-inflammatory cytokines from activated neutrophils. This selective modulation - enhanced killing with reduced inflammation - represents an ideal antimicrobial response that maximizes pathogen clearance while minimizing collateral tissue damage (Zheng et al., 2007).
LL-37 also influences neutrophil lifespan. Under normal circumstances, neutrophils undergo apoptosis (programmed cell death) within 24 hours of release from the bone marrow, a process essential for limiting inflammation. LL-37 can either promote or delay neutrophil apoptosis depending on the concentration and environmental context. At low concentrations, LL-37 tends to promote neutrophil survival through anti-apoptotic signaling via PI3K/Akt and ERK1/2 pathways, extending their active antimicrobial function. At higher concentrations, LL-37 can induce secondary necrosis in neutrophils, releasing their intracellular contents (including more LL-37) and creating a self-amplifying antimicrobial cascade.
NETosis - neutrophil extracellular trap formation - is another process modulated by LL-37. When neutrophils undergo NETosis, they extrude their nuclear DNA along with antimicrobial proteins to create web-like structures (NETs) that trap and kill bacteria. LL-37 promotes NET formation and is itself a component of NETs, where its antimicrobial activity contributes to pathogen killing within the NET structure. However, excessive NET formation has been implicated in several inflammatory and autoimmune conditions, creating another context where LL-37's immune-activating properties can become pathological.
Macrophage Modulation
Macrophages are professional phagocytes that serve as both sentinels of the innate immune system and bridges to adaptive immunity. LL-37 modulates macrophage function at multiple levels, from initial pathogen detection to cytokine production and antigen presentation.
One of the most significant effects of LL-37 on macrophages is the modulation of Toll-like receptor signaling. LL-37 can both enhance and suppress TLR-mediated responses depending on the specific TLR pathway involved. For TLR2 (which detects Gram-positive bacterial products) and TLR4 (which detects LPS), LL-37 generally acts as a modifier that fine-tunes the response rather than simply amplifying or suppressing it. At the molecular level, this involves differential effects on NF-kappaB and MAPK signaling cascades downstream of TLR activation.
In the context of LPS stimulation, LL-37 reduces macrophage production of TNF-alpha, IL-12, and nitric oxide while maintaining or enhancing IL-10 production. This shifts the cytokine profile from a strongly pro-inflammatory pattern toward a more balanced response that supports pathogen clearance without excessive tissue-damaging inflammation. The mechanism involves LL-37 binding to LPS and altering how it is presented to the TLR4/MD-2 complex, resulting in modified signaling through MyD88-dependent and TRIF-dependent pathways.
LL-37 also promotes macrophage polarization toward the M2 phenotype, which is associated with tissue repair and anti-inflammatory functions. M2 macrophages produce growth factors (TGF-beta, VEGF, PDGF) that support wound healing, promote angiogenesis, and facilitate extracellular matrix remodeling. This effect of LL-37 on macrophage polarization provides another mechanism through which the peptide supports wound repair beyond its direct effects on keratinocytes and fibroblasts.
In tuberculosis, LL-37's effects on macrophages are critically important. The peptide is induced by vitamin D-dependent signaling in M. tuberculosis-infected macrophages and then directed to mycobacteria-containing phagosomes, where it participates in intracellular killing. LL-37 also promotes autophagy in macrophages, a cellular self-digestion process that is one of the most effective mechanisms for eliminating intracellular M. tuberculosis. The vitamin D-cathelicidin-autophagy axis represents one of the most elegant examples of innate immune defense against intracellular pathogens.
Dendritic Cell Modulation
Dendritic cells (DCs) are the most potent antigen-presenting cells in the immune system, serving as the primary link between innate pathogen detection and adaptive immune activation. LL-37 influences dendritic cell biology at multiple stages, from differentiation through maturation to antigen presentation.
During DC differentiation from monocyte precursors, LL-37 promotes the development of DCs with enhanced antigen-capturing capacity and increased expression of co-stimulatory molecules (CD80, CD86, CD40). These LL-37-conditioned DCs are more efficient at stimulating T cell proliferation and cytokine production, leading to stronger adaptive immune responses against encountered pathogens.
LL-37 also facilitates antigen delivery to DCs. The peptide can complex with microbial products (DNA, RNA, proteins) and enhance their uptake by DCs through receptor-mediated endocytosis. This "chaperoning" function means that LL-37 not only kills pathogens but also ensures that their antigenic material is efficiently delivered to the antigen-presenting machinery, optimizing the development of adaptive immunity.
The DC-modulating effects of LL-37 have implications for vaccine design. Researchers have explored using LL-37 as an immunoadjuvant - a substance that enhances vaccine immune responses. When co-administered with antigens, LL-37 enhances DC maturation and antigen presentation, leading to stronger antibody and T cell responses. This adjuvant activity could potentially improve the efficacy of vaccines against pathogens where current vaccines are suboptimal.
Mast Cell Interactions
Mast cells reside in tissues throughout the body, particularly at barrier surfaces like the skin, lungs, and gut, where they serve as sentinel cells that detect pathogens and initiate inflammatory responses. LL-37 has potent effects on mast cells that contribute to both its antimicrobial and its wound healing functions.
LL-37 is a potent mast cell degranulator, triggering the release of histamine, tryptase, and other preformed mediators from mast cell granules. This degranulation response occurs through a mechanism involving Mas-related G protein-coupled receptor X2 (MRGPRX2), a receptor that mast cells use to detect cationic molecules in their environment. The resulting histamine release increases local blood flow and vascular permeability, facilitating the influx of additional immune cells from the bloodstream.
Beyond degranulation, LL-37 stimulates mast cell production of several cytokines including IL-1beta, IL-4, IL-5, IL-6, and IL-31. These cytokines shape the local immune environment and influence the recruitment and activation of other immune cell types. The IL-4 and IL-5 production, in particular, contributes to Th2-type immune responses that may be relevant in parasitic infection defense and allergic inflammation.
The mast cell-activating properties of LL-37 also explain one of its most common side effects: injection site reactions. The burning, redness, and swelling reported at subcutaneous injection sites are consistent with local mast cell degranulation and histamine release. While generally mild and self-limiting, these reactions can be minimized by using lower concentrations, injecting slowly, and rotating injection sites.
T Cell and Adaptive Immunity Effects
While LL-37 is primarily an innate immune molecule, it has significant effects on adaptive immunity, both directly through interactions with T cells and indirectly through its effects on dendritic cells and other antigen-presenting cells.
LL-37 is chemotactic for T cells, attracting them to sites of infection through FPRL-1 (FPR2/ALX) receptor signaling. This chemotactic activity operates at nanomolar concentrations, well below the levels needed for direct antimicrobial killing, meaning that even trace amounts of LL-37 can begin recruiting T cells to infected tissues.
The peptide also influences T cell differentiation and function. Through its effects on dendritic cell cytokine production, LL-37 tends to promote Th1-polarized T cell responses, which are optimal for defense against intracellular pathogens (viruses, mycobacteria, certain parasites). This Th1-promoting effect is consistent with LL-37's role as an anti-infectious agent and distinguishes it from some other antimicrobial peptides that promote Th2 responses.
Recent research has identified a potential role for LL-37 in regulatory T cell (Treg) biology. LL-37 may influence the balance between effector T cells and Tregs, potentially affecting immune tolerance and autoimmune risk. This area of research is still in its early stages, but it has implications for understanding the autoimmune conditions (psoriasis, lupus) in which LL-37 has been implicated.
LL-37 in Sepsis and Critical Care
Sepsis Pathophysiology and LL-37
Sepsis is a life-threatening condition caused by the body's dysregulated response to infection, affecting approximately 49 million people worldwide each year and causing an estimated 11 million deaths. The condition involves a complex interplay between pathogen burden, endotoxin release, cytokine storm, and multi-organ dysfunction. LL-37's dual antimicrobial and immunomodulatory properties position it as a potentially valuable therapeutic agent in this context.
The endotoxin-neutralizing capacity of LL-37 is particularly relevant in Gram-negative sepsis. During antibiotic treatment of Gram-negative infections, bacterial lysis releases large quantities of LPS, which can paradoxically worsen the clinical picture by triggering massive inflammatory activation - the so-called Jarisch-Herxheimer reaction. LL-37's ability to bind and neutralize free LPS could potentially mitigate this complication, providing a rationale for combining LL-37 with conventional antibiotics in sepsis management.
Animal studies support this concept. In murine sepsis models (cecal ligation and puncture), administration of LL-37 or the murine cathelicidin CRAMP improved survival rates by 25-40%, reduced circulating endotoxin and cytokine levels, attenuated organ damage markers, and improved bacterial clearance from the bloodstream. These effects were observed even when cathelicidin was administered several hours after the onset of sepsis, suggesting a therapeutic window that could be clinically applicable (Nagaoka et al., 2020).
However, the translation of these animal findings to human sepsis is complicated by several factors. The murine sepsis model uses CRAMP (the murine cathelicidin) rather than LL-37, and species differences in cathelicidin biology, particularly the absence of the vitamin D regulatory axis in mice, limit the direct applicability of murine results. Additionally, human sepsis is far more heterogeneous than experimental sepsis models, with varying causative organisms, patient comorbidities, and immune responses making one-size-fits-all therapeutic approaches challenging.
Circulating LL-37 as a Biomarker
Several studies have investigated circulating LL-37 levels as a biomarker for infection severity and sepsis outcome. Results have been mixed. Some studies show that high LL-37 levels at the time of infection diagnosis correlate with better outcomes, suggesting that strong cathelicidin production reflects an effective innate immune response. Other studies have found elevated LL-37 levels in patients with poor outcomes, potentially reflecting excessive inflammation or massive neutrophil activation and death.
These contradictory findings likely reflect the complex, dual-function nature of LL-37. In early infection, high LL-37 levels may indicate a strong antimicrobial response that limits pathogen spread. In late-stage sepsis, high LL-37 levels may indicate massive tissue damage, neutrophil NET formation, and loss of immune homeostasis. The timing and context of measurement are therefore critical for interpreting LL-37 levels as biomarkers.
Despite these complexities, LL-37 measurement may have value as part of a multi-marker panel for sepsis diagnosis and prognosis, combined with procalcitonin, C-reactive protein, and other established infection biomarkers. The development of rapid point-of-care LL-37 assays could facilitate clinical implementation of such panels.
Neonatal and Pediatric Considerations
LL-37 in Neonatal Immunity
Neonates are particularly vulnerable to infections due to the immaturity of both their innate and adaptive immune systems. LL-37 plays a crucial role in neonatal antimicrobial defense, and understanding its expression patterns in early life has important implications for neonatal health.
Cord blood LL-37 levels are generally lower than adult levels, reflecting the relative immaturity of neonatal innate immunity. However, LL-37 is present in significant quantities in the vernix caseosa, the waxy coating that covers newborns at birth. The vernix provides a physical and chemical barrier that protects the newborn's skin during the transition from the sterile intrauterine environment to the microbe-rich extrauterine world. LL-37 in the vernix contributes to this protective function by providing broad-spectrum antimicrobial activity on the skin surface during the first days of life.
Breast milk is another important source of LL-37 for neonates. Human breast milk contains detectable levels of LL-37, which may contribute to the well-documented protective effects of breastfeeding against neonatal infections. The peptide is present in colostrum at higher concentrations than in mature milk, consistent with a role in protecting the newborn during the period of greatest vulnerability.
The vitamin D-LL-37 axis is operational in neonatal immune cells. Mandic Havelka et al. (2010) demonstrated that vitamin D3 induces CAMP gene expression in newborn blood cells, confirming that the molecular machinery for vitamin D-mediated cathelicidin induction is present at birth. This finding supports the practice of vitamin D supplementation in breastfed infants, which may serve not only bone health but also antimicrobial defense through enhanced LL-37 production.
Premature infants face an even greater infection risk and show lower LL-37 levels compared to full-term neonates. The deficiency is compounded by the premature infant's thin, easily damaged skin, immature gut barrier, and reduced capacity for inflammatory signaling. Research into strategies for boosting LL-37 levels in premature infants, including vitamin D supplementation and topical LL-37 application, is an active area of investigation in neonatal medicine.
Pediatric Infection Susceptibility
In older children, LL-37 levels gradually increase toward adult values as the immune system matures. However, certain pediatric populations show persistently low LL-37 levels that may contribute to infection susceptibility. Children with atopic dermatitis, as discussed earlier, have reduced skin LL-37 expression that predisposes to bacterial skin infections. Children with cystic fibrosis show compromised LL-37 activity in the airways due to the high-salt environment. And children with vitamin D deficiency, which is common in northern latitudes, may have globally reduced LL-37 production.
The relationship between vitamin D status, LL-37 levels, and pediatric respiratory infections has been studied in several clinical trials. While results have been inconsistent, meta-analyses suggest that vitamin D supplementation reduces the risk of acute respiratory infections in children, with the strongest effects seen in children who are vitamin D deficient at baseline. The LL-37-mediated enhancement of antimicrobial defense is thought to be a major mechanism underlying this protective effect.
LL-37 in Urinary Tract and Reproductive Health
Urinary Tract Defense
The urinary tract epithelium produces LL-37 as part of its antimicrobial defense system. Urothelial cells in the bladder and ureter express CAMP and produce LL-37, which is secreted into the urine where it contributes to the antimicrobial properties of this body fluid. Urinary LL-37 concentrations increase significantly during urinary tract infections (UTIs), reflecting both increased urothelial production and neutrophil infiltration into the urinary tract.
Uropathogenic E. coli (UPEC), the most common cause of UTIs, is susceptible to LL-37 at MIC values of 1-4 micrograms/mL. However, the effectiveness of urinary LL-37 is influenced by urine composition, particularly pH and osmolality. Acidic urine pH can reduce LL-37 activity, while the high osmolality of concentrated urine may interfere with peptide-membrane interactions. These factors help explain why adequate hydration, which produces more dilute urine, is associated with reduced UTI risk.
Recurrent UTIs, which affect approximately 25% of women who experience an initial UTI, may involve deficient LL-37 production. Studies have found lower urothelial CAMP expression in women prone to recurrent UTIs compared to those without recurrence, suggesting that impaired local antimicrobial peptide production contributes to susceptibility. Vitamin D supplementation has been explored as a strategy to boost urinary LL-37 levels and reduce UTI recurrence, with some clinical studies showing promising results.
Reproductive Tract Defense
LL-37 is expressed throughout the female reproductive tract, including the vaginal epithelium, cervix, endometrium, and fallopian tubes. In the vaginal environment, LL-37 contributes to defense against sexually transmitted pathogens including Neisseria gonorrhoeae, Chlamydia trachomatis, and Candida species. The peptide also shows activity against bacterial vaginosis-associated organisms.
In the male reproductive tract, LL-37 is present in seminal fluid at concentrations of approximately 1-5 micrograms/mL. It contributes to the antimicrobial properties of semen and may play a role in protecting against sexually transmitted infections. Some studies have also suggested a role for LL-37 in sperm function and fertility, though this area remains poorly characterized.
The reproductive tract expression of LL-37 is regulated by sex hormones in addition to vitamin D. Estrogen upregulates CAMP expression in vaginal and cervical epithelial cells, which may contribute to the cyclical variation in vaginal antimicrobial defense observed across the menstrual cycle. This hormonal regulation also explains the increased susceptibility to vaginal infections seen during periods of relative estrogen deficiency, such as menopause.
Manufacturing and Synthesis Approaches
Chemical Synthesis
LL-37 is commercially produced primarily through solid-phase peptide synthesis (SPPS), a chemical manufacturing process that builds the peptide one amino acid at a time on a solid support resin. The 37-amino-acid length of LL-37 is at the upper end of what can be efficiently produced by SPPS, and the synthesis becomes technically challenging due to the peptide's tendency to aggregate during chain assembly.
The synthesis process involves repeated cycles of amino acid coupling (adding one residue at a time), deprotection (removing the chemical protecting group from the growing chain), and washing. After all 37 residues have been assembled, the peptide is cleaved from the resin and purified by preparative HPLC. The overall yield from a single synthesis run is typically 20-40%, with losses at each coupling step accumulating to reduce the final yield.
The cost of synthetic LL-37 production is substantial, primarily due to the large number of coupling cycles required, the expensive amino acid building blocks, and the purification requirements. Research-grade LL-37 typically costs $50-200 per milligram depending on purity and quantity, while pharmaceutical-grade material suitable for clinical use would cost significantly more due to stringent quality control requirements, GMP manufacturing standards, and regulatory compliance testing.
Recombinant Production
To address the high cost of chemical synthesis, researchers have developed recombinant production methods for LL-37 using bacterial expression systems. E. coli is the most commonly used host organism, with the CAMP gene (or an optimized synthetic gene) expressed as a fusion protein to protect the host cell from LL-37's antimicrobial activity. After expression, the fusion partner is cleaved and the LL-37 peptide is purified.
Recombinant production offers potential cost advantages for large-scale manufacturing, with estimated production costs 5-10 fold lower than chemical synthesis. However, recombinant LL-37 requires careful quality control to ensure the absence of host cell proteins, endotoxin contamination (a particular concern with E. coli expression systems), and post-translational modifications that could affect activity or immunogenicity.
Alternative expression systems, including yeast (Pichia pastoris), insect cells, and plant cells, have been explored for recombinant LL-37 production. Each system offers advantages and limitations in terms of yield, purity, scalability, and cost. The choice of production system is an active area of development that will influence the commercial viability of LL-37-based therapeutics.
Rational Analog Design
The limitations of native LL-37 (proteolytic instability, potential toxicity at high concentrations, high production cost) have driven extensive efforts to design improved analogs. These analogs are typically shorter than the full-length 37-amino-acid peptide, retaining the minimum structural elements needed for activity while eliminating regions that contribute to instability or toxicity.
The shortest active fragment of LL-37, designated KR-12 (residues 18-29, just 12 amino acids), retains antimicrobial activity but with reduced potency compared to the full-length peptide. Intermediate fragments like FK-16 (residues 17-32) and GF-17 (residues 17-32) offer a better balance of activity, stability, and manufacturing cost. These shorter analogs can be produced by SPPS at significantly lower cost than full-length LL-37, making them more commercially viable for therapeutic development.
Beyond simple truncation, researchers have employed several strategies to improve LL-37 analog properties. D-amino acid substitution at protease-sensitive sites dramatically increases plasma stability without necessarily reducing antimicrobial activity. Cyclization of the peptide backbone restricts conformational flexibility and can improve both stability and selectivity. Lipidation (attaching fatty acid chains to the peptide) can enhance membrane interaction and provide sustained-release properties. PEGylation (conjugation with polyethylene glycol) extends circulating half-life by reducing renal clearance and proteolytic degradation.
The analog P60.4Ac, derived from LL-37, has emerged as one of the most promising modified cathelicidin peptides. It retains broad-spectrum antimicrobial and anti-biofilm activity, shows improved stability compared to native LL-37, and demonstrates reduced cytotoxicity toward mammalian cells. Clinical development of P60.4Ac and similar analogs is anticipated to proceed more rapidly than development of the native LL-37 peptide, given their improved therapeutic profiles.

LL-37 in Medical Device and Implant Applications
The Problem of Device-Associated Infections
Medical device-associated infections represent one of the most significant challenges in modern healthcare. Central venous catheters, urinary catheters, orthopedic implants, cardiac devices, and dental implants all provide surfaces where bacterial biofilms can establish, leading to persistent infections that are extremely difficult to treat. An estimated 1-2% of all orthopedic implants become infected, requiring revision surgery in many cases. Central line-associated bloodstream infections (CLABSIs) affect approximately 250,000 patients annually in the United States alone.
The fundamental problem is that bacteria adhere to implant surfaces within hours of implantation, forming biofilms that resist both antibiotic treatment and immune clearance. The biofilm bacteria exist in a metabolically dormant state (persister cells) that makes them virtually invulnerable to conventional antibiotics, most of which target actively growing cells. Even after apparently successful antibiotic treatment, persister cells can reactivate and re-establish infection when therapy is discontinued.
LL-37's combination of anti-biofilm and antimicrobial properties makes it a compelling candidate for preventing and treating device-associated infections. Unlike conventional antibiotics, LL-37's membrane-disrupting mechanism is effective against both actively growing and dormant bacteria, potentially addressing the persister cell problem that limits antibiotic efficacy against biofilm infections.
Surface Coating Strategies
Researchers have developed several approaches for incorporating LL-37 into medical device surfaces. Covalent immobilization involves chemically bonding LL-37 to the device surface through linker molecules, creating a permanent antimicrobial coating that resists leaching. Physical adsorption uses electrostatic or hydrophobic interactions to attach LL-37 to surfaces without chemical modification, offering simpler manufacturing but potentially less durable coatings.
Layer-by-layer (LbL) assembly is a particularly versatile approach that builds alternating layers of LL-37 and an oppositely charged polymer on the device surface. This technique allows precise control over the amount of peptide loaded and can be designed to provide either sustained release or a permanent contact-killing surface. LbL coatings containing LL-37 have shown excellent anti-biofilm activity against S. aureus and P. aeruginosa in in vitro studies.
Hydroxyapatite coatings, commonly used on orthopedic implants to promote bone integration, can be combined with LL-37 to create dual-function surfaces that both resist infection and promote osseointegration. The peptide can be adsorbed onto or incorporated within the hydroxyapatite layer, providing antimicrobial activity during the critical post-implantation period when the risk of infection is highest.
Titanium dental implants coated with LL-37 have been tested in laboratory models of peri-implantitis. The LL-37-coated surfaces showed significantly reduced bacterial adhesion and biofilm formation compared to uncoated titanium, while maintaining compatibility with osteoblast cells needed for implant integration. These results suggest that LL-37 coatings could help prevent peri-implantitis, which affects 20-40% of dental implant recipients and is the leading cause of implant failure.
Catheter Applications
Catheter-associated urinary tract infections (CAUTIs) are the most common type of healthcare-associated infection, accounting for approximately 40% of all nosocomial infections. LL-37-coated urinary catheters have been studied as a strategy to prevent bacterial colonization and biofilm formation on catheter surfaces.
In vitro studies have demonstrated that LL-37-coated catheters resist colonization by common CAUTI pathogens including E. coli, Enterococcus faecalis, Klebsiella pneumoniae, and Candida albicans. The anti-biofilm activity of the coating persisted for several days to weeks depending on the coating technology used, with controlled-release formulations showing the longest duration of activity.
Central venous catheter applications have also been explored. CLABSIs carry a mortality rate of approximately 12-25% and add an estimated $25,000-$45,000 to the cost of a hospital stay. LL-37-modified catheter surfaces have shown promising anti-bacterial and anti-biofilm properties in preclinical testing, though no clinical trials of LL-37-coated catheters have been completed to date.
Wound Dressing Integration
LL-37 has been incorporated into various wound dressing materials to create antimicrobial dressings for chronic wound management. Electrospun nanofiber dressings loaded with LL-37 provide high surface area for sustained peptide release while creating a physical barrier that protects the wound from external contamination. Hydrogel dressings containing LL-37 maintain a moist wound environment (optimal for healing) while providing continuous antimicrobial activity.
A particularly innovative approach combines LL-37 with thermoresponsive polymers that undergo a sol-gel transition at body temperature. The peptide-loaded solution is applied to the wound as a liquid, then gels upon contact with the warm wound bed, forming a conformal antimicrobial coating that adheres to the irregular wound surface and provides sustained LL-37 release over several days.
These advanced wound dressing technologies address a key limitation of topical LL-37 therapy: the peptide's rapid degradation by wound fluid proteases. By encapsulating or binding LL-37 within protective matrices, these formulations extend the duration of antimicrobial activity from hours (for a simple LL-37 solution) to days or weeks, reducing the frequency of dressing changes and improving patient comfort.
LL-37 in the Context of the Antibiotic Resistance Crisis
The Global Antibiotic Resistance Challenge
Antibiotic resistance has been declared one of the greatest threats to global public health by the World Health Organization. An estimated 1.27 million deaths were directly attributable to antibiotic-resistant infections in 2019, making antimicrobial resistance a leading cause of death worldwide. Without effective interventions, projections suggest that antibiotic-resistant infections could cause 10 million deaths annually by 2050, surpassing cancer as a cause of mortality.
The development pipeline for new antibiotics has not kept pace with the evolution of resistance. Most major pharmaceutical companies have withdrawn from antibiotic development due to unfavorable economics - the return on investment for new antibiotics (which are used for short courses) cannot compete with drugs for chronic conditions. This market failure has left a growing gap between the antibiotics available and the resistant organisms that need to be treated.
Antimicrobial peptides like LL-37 represent a fundamentally different approach to anti-infective therapy that could help address this crisis. Their membrane-active mechanisms are inherently less susceptible to conventional resistance mechanisms, their multi-target mode of action creates a higher barrier to resistance development, and their immunomodulatory properties provide added therapeutic value beyond direct pathogen killing.
Why LL-37 Resistance Is Difficult to Evolve
The evolution of high-level resistance to LL-37 faces several fundamental barriers. First, the primary target is the cell membrane itself, not a specific protein or enzyme that can be altered by a single gene mutation. Bacteria would need to fundamentally restructure their membrane lipid composition to resist LL-37, which would compromise essential membrane functions like nutrient transport, energy production, and cell signaling.
Second, LL-37 attacks multiple intracellular targets in addition to the membrane. Even if a bacterium could partially protect its membrane, it would still need to prevent LL-37 from binding to DNA, ribosomes, and cell wall biosynthetic enzymes. Simultaneous evolution of resistance at all these targets is extremely unlikely through spontaneous mutation.
Third, LL-37 operates at concentrations (micrograms per milliliter) that are orders of magnitude higher than the concentrations of most antibiotics (nanograms to micrograms per milliliter). This means that the selective pressure for resistance is applied at concentrations well above the minimum inhibitory level, leaving little opportunity for bacteria with marginal resistance to survive and accumulate additional resistance mechanisms.
Experimental evolution studies have confirmed these theoretical predictions. When bacteria are exposed to gradually increasing concentrations of LL-37 over many generations, they develop at most modest (2-4 fold) increases in MIC, compared to the 100-1000 fold increases commonly seen with serial antibiotic exposure. This limited capacity for resistance evolution makes LL-37 and similar antimicrobial peptides attractive options for therapeutic development.
LL-37 as a Complement to Conventional Antibiotics
Rather than replacing conventional antibiotics, LL-37 is most likely to find clinical utility as a complement to existing antibiotic therapy. Several combination strategies have been proposed:
Sensitizer strategy: Using sub-MIC concentrations of LL-37 to permeabilize bacterial membranes, increasing the intracellular concentration of co-administered antibiotics. This approach has been shown to restore susceptibility of MRSA strains to beta-lactam antibiotics in vitro, potentially reviving entire antibiotic classes that have been rendered ineffective by resistance.
Anti-biofilm adjunct: Using LL-37 to disrupt biofilms that protect bacteria from both antibiotics and immune clearance, making the embedded bacteria accessible to conventional antibiotic therapy. This is particularly relevant for chronic infections where biofilm formation is the primary barrier to treatment success.
Endotoxin shield: Using LL-37 concurrently with antibiotics for Gram-negative infections to neutralize the LPS released by antibiotic-induced bacterial lysis, preventing the endotoxin-mediated inflammatory surge that can worsen clinical outcomes in sepsis.
Immune system activator: Using LL-37's immunomodulatory properties to enhance the patient's own immune response, working cooperatively with antibiotics to achieve faster pathogen clearance and reduce the duration of antibiotic therapy needed. Shorter antibiotic courses reduce the selective pressure for resistance development and decrease the risk of antibiotic side effects including Clostridioides difficile infection.
Each of these strategies represents an opportunity to extend the useful life of existing antibiotics while addressing the biofilm and resistance challenges that limit current antibiotic efficacy. The development of LL-37 or its analogs for these combination applications could have a significant impact on the antibiotic resistance crisis, even if the peptides are never used as standalone antimicrobial agents.
Regulatory Landscape and Development Status
Current Regulatory Status
LL-37 is not currently approved by the FDA or any other regulatory agency for any therapeutic indication. It is available as a research peptide and is classified as an investigational compound. The regulatory pathway for LL-37 as a therapeutic agent would depend on the specific application: topical formulations for wound healing would follow the new drug application (NDA) pathway, medical device coatings would require device-specific regulatory approvals, and injectable formulations would face the most stringent regulatory requirements.
The lack of patent protection for the native LL-37 sequence (it is a naturally occurring human peptide) has complicated commercial development. Without strong patent protection, pharmaceutical companies have limited economic incentive to invest in the expensive clinical trials needed for regulatory approval. This "natural molecule" problem is a common barrier for antimicrobial peptide therapeutics and has led most commercial development efforts to focus on patentable synthetic analogs rather than the native peptide.
Several LL-37-derived analogs are in various stages of preclinical and early clinical development. The analog P60.4Ac has been studied in clinical settings for ear nose and throat applications. Other analogs are in preclinical development for wound healing, respiratory infection, and medical device coating applications. The timeline from current development stage to potential market availability is estimated at 5-10 years for the most advanced candidates.
Research Access and Availability
For researchers and clinicians, LL-37 is currently available through peptide synthesis companies and specialized peptide suppliers. Research-grade LL-37 is available in quantities ranging from milligrams to grams, with prices that have decreased significantly over the past decade as synthesis technology has improved.
FormBlends LL-37 provides research-grade peptide with documented purity specifications and quality testing. For individuals interested in exploring LL-37 as part of a supervised research protocol, the Free Assessment can help determine appropriate approaches, and the Dosing Calculator provides tools for accurate reconstitution and dosing calculations.
LL-37 in Ophthalmic and Ocular Health
Ocular Surface Antimicrobial Defense
The ocular surface is constantly exposed to environmental microorganisms yet rarely develops clinically significant infections in healthy individuals. This remarkable resilience is due in part to the antimicrobial peptides present in tears and the ocular surface epithelium, with LL-37 playing a prominent role in this defense system.
LL-37 is expressed by the corneal and conjunctival epithelia and is present in tear fluid at concentrations of approximately 0.5-3 micrograms per milliliter. The peptide is produced constitutively by surface epithelial cells and can be upregulated in response to infection or injury. In addition to direct antimicrobial activity, LL-37 in the tear film contributes to wound healing after corneal injuries - abrasions, ulcerations, and post-surgical wounds all benefit from LL-37's combined antimicrobial and regenerative properties.
Research by Gordon et al. (2005) demonstrated that human corneal and conjunctival epithelial cells express CAMP and produce LL-37 with antimicrobial activity against common ocular pathogens including Staphylococcus aureus, Pseudomonas aeruginosa, and Streptococcus pneumoniae. The expression was upregulated by exposure to bacterial products, confirming a responsive defense system that scales its output based on microbial challenge.
Contact lens wear significantly alters the ocular surface antimicrobial environment. Contact lenses can disrupt the tear film, reduce oxygen delivery to the cornea, and provide a surface for bacterial adhesion and biofilm formation. Studies have shown that contact lens wearers have altered tear LL-37 levels, which may contribute to the increased risk of microbial keratitis associated with contact lens use. Understanding these dynamics could lead to strategies for reducing contact lens-related infections, potentially including LL-37-modified contact lens surfaces or LL-37-containing lens care solutions.
Corneal Infections and Keratitis
Microbial keratitis (corneal infection) is a sight-threatening condition that requires urgent treatment. P. aeruginosa is the most common cause of contact lens-related keratitis and one of the most destructive corneal pathogens. S. aureus, Streptococcus pneumoniae, and Fusarium species are also significant causes. LL-37 shows activity against all of these organisms, positioning it as a potential topical therapeutic for corneal infections.
The ocular application of LL-37 presents unique pharmacological challenges. The rapid turnover of the tear film (approximately every 5-7 minutes) means that topically applied LL-37 would be quickly diluted and washed away. Sustained-release formulations, such as LL-37-loaded nanoparticles or contact lenses that slowly release the peptide, could overcome this limitation by providing continuous antimicrobial activity on the ocular surface.
In Acanthamoeba keratitis, a particularly difficult-to-treat corneal infection caused by free-living amoebae, LL-37 has shown activity against both the trophozoite (active) and cyst (dormant) forms of the organism. This is clinically significant because Acanthamoeba cysts are extremely resistant to conventional antimicrobial agents, and their persistence is a major reason for treatment failure and disease recurrence. LL-37's ability to damage cyst walls through its membrane-disrupting mechanism offers a potential advantage over current Acanthamoeba therapies.
LL-37 Drug Development Pipeline and Future Therapeutics
Current Development Pipeline
Several therapeutic programs based on LL-37 or cathelicidin-derived peptides are in various stages of development. While none have reached late-stage clinical trials for the native LL-37 molecule, the breadth of preclinical activity and early clinical safety data have maintained interest from both academic researchers and smaller biotechnology companies.
The most advanced clinical program involves topical LL-37 for chronic wound healing, building on the positive phase I/II results of Gronberg et al. (2014). Additional clinical studies are planned or underway to extend these findings to other wound types and to optimize dosing regimens. The wound healing application benefits from the topical delivery route, which avoids the proteolytic stability challenges of systemic administration and achieves high local concentrations at the target site.
The analog P60.4Ac (also known as OP-145) has been evaluated in clinical settings for middle ear infections (chronic suppurative otitis media). This application leverages the peptide's antimicrobial and anti-biofilm properties in a confined anatomical space where high local concentrations can be achieved through direct instillation. Early clinical results have been described as promising, though detailed published data are limited.
Several academic groups are developing LL-37-based approaches for respiratory infections, including inhaled formulations for lung delivery. The rationale is compelling: the respiratory epithelium is a natural production site for LL-37, inhalation bypasses systemic stability challenges, and the local immune-modulating effects of LL-37 could provide both antimicrobial and anti-inflammatory benefits in respiratory infections. Development is at the preclinical stage, with proof-of-concept studies in animal models of influenza, RSV, and bacterial pneumonia.
Combination Product Development
The cooperative interactions between LL-37 and conventional antibiotics have led to interest in combination products that package both agents together. A catheter coated with both LL-37 and a conventional antibiotic, for example, could provide enhanced anti-biofilm protection through dual mechanisms. Similarly, wound dressings containing LL-37 combined with silver nanoparticles or conventional antibiotics could offer broader antimicrobial coverage than either component alone.
The development of LL-37-antibiotic combination products faces regulatory complexity, as these products may need to satisfy requirements for both drug approval and device approval. However, the potential clinical benefits of such combinations - particularly for preventing device-associated infections - justify the additional regulatory burden.
Gene Therapy and Endogenous Production Enhancement
Rather than administering exogenous LL-37, an alternative therapeutic strategy involves enhancing the body's own LL-37 production. Vitamin D supplementation is the simplest approach, as discussed earlier. But more advanced strategies are also being explored.
Gene therapy approaches involve delivering the CAMP gene to target tissues using viral or non-viral vectors, enabling sustained local production of LL-37 by the patient's own cells. Adeno-associated virus (AAV) vectors carrying the CAMP gene have been tested in animal models of skin wound healing, showing enhanced wound closure and reduced bacterial burden at the wound site. The advantage of gene therapy over exogenous peptide delivery is the potential for sustained, self-renewing production without repeated dosing.
Another approach involves pharmacological induction of endogenous CAMP expression. Beyond vitamin D, several small molecules have been identified that can upregulate CAMP gene transcription. Butyrate and other HDAC inhibitors, certain bile acids acting through VDR or FXR, and specific phenylbutyrate derivatives have all shown CAMP-inducing activity. The development of orally bioavailable small molecules that selectively induce cathelicidin expression could provide a convenient, cost-effective approach to boosting antimicrobial peptide defenses.
Probiotics that enhance endogenous LL-37 production represent yet another strategy. Certain Lactobacillus and Bifidobacterium strains have been shown to upregulate CAMP expression in intestinal epithelial cells, suggesting that targeted probiotic supplementation could enhance gut mucosal LL-37 production. This approach is particularly attractive because it leverages the existing gut microbiome-epithelium interaction to boost local antimicrobial defense without the need for peptide synthesis or delivery.
Development Outlook
The LL-37 therapeutic development landscape includes multiple approaches: native peptide formulations for topical use, optimized synthetic analogs for broader applications, medical device coatings, combination products with conventional antibiotics, gene therapy for sustained production, and pharmacological induction of endogenous expression. This diversity of approaches increases the likelihood that at least some LL-37-based therapeutics will reach clinical use, even if the specific pathway to market differs from the traditional single-agent drug development model. The Science & Research section at FormBlends tracks developments in cathelicidin-based therapeutics as they progress through the development pipeline.
Emerging Combination Therapies and Novel Approaches
Photobiomodulation and LL-37
Photobiomodulation (PBM), also known as low-level laser therapy (LLLT), involves the application of red or near-infrared light to biological tissues. PBM has been shown to enhance wound healing, reduce inflammation, and promote tissue repair through mechanisms involving mitochondrial chromophore activation and downstream signaling cascades. Recent research has explored the intersection between PBM and antimicrobial peptide biology, revealing that light therapy can upregulate LL-37 expression in skin and mucosal tissues.
The mechanism appears to involve PBM-induced activation of the vitamin D receptor and enhanced intracellular vitamin D metabolism. Red and near-infrared light can stimulate the expression of CYP27B1, the enzyme that converts 25-hydroxyvitamin D to the active 1,25-dihydroxyvitamin D3 form in keratinocytes and other cell types. This locally increased active vitamin D then drives CAMP gene transcription through the established vitamin D response element. The result is enhanced LL-37 production in light-exposed tissues.
This finding has practical implications for wound management. Combining PBM with topical LL-37 application could provide combined benefits - the exogenous LL-37 provides immediate antimicrobial protection while the PBM-stimulated endogenous production provides sustained antimicrobial defense. Clinical studies investigating this combination approach for chronic wound healing are in the planning stages at several academic medical centers.
Antimicrobial Photodynamic Therapy Combinations
Antimicrobial photodynamic therapy (aPDT) uses a photosensitizer molecule activated by visible light to generate reactive oxygen species (ROS) that kill bacteria. When combined with LL-37, aPDT shows enhanced antimicrobial activity compared to either approach alone. The cooperative enhancement arises because LL-37's membrane-disrupting activity increases bacterial permeability to photosensitizer molecules, allowing greater intracellular accumulation and more potent light-activated killing.
This combination is particularly promising for biofilm infections, where both LL-37 and aPDT face challenges when used individually. LL-37 can penetrate and disrupt the biofilm matrix, exposing embedded bacteria to the photosensitizer, while the ROS generated by aPDT can damage bacteria that have partially resisted LL-37's membrane effects. The dual mechanism attack leaves fewer viable bacteria and reduces the risk of treatment failure.
Probiotic Synergies
The intersection of probiotics and antimicrobial peptides represents a growing research area with significant therapeutic potential. Certain probiotic strains produce their own antimicrobial peptides (bacteriocins) that can work cooperatively with human LL-37 to provide enhanced pathogen suppression in the gut and on skin surfaces.
Lactobacillus rhamnosus GG, one of the most extensively studied probiotic strains, has been shown to upregulate CAMP gene expression in intestinal epithelial cells through a mechanism involving the Lactobacillus surface protein p40 and downstream EGFR signaling. This means that probiotic supplementation with appropriate strains can boost the body's own LL-37 production in the gut, providing enhanced mucosal defense without the need for exogenous peptide administration.
The practical implication is that probiotic supplementation could serve as a low-cost, low-risk strategy for enhancing endogenous LL-37 production, particularly in the gut. Combined with adequate vitamin D status and a fiber-rich diet that supports butyrate production, probiotics could create optimal conditions for maximal endogenous cathelicidin expression. This "nutritional and microbial optimization" approach represents the most accessible entry point for individuals seeking to enhance their natural antimicrobial peptide defenses.
Cold Atmospheric Plasma Therapy
Cold atmospheric plasma (CAP) therapy is an emerging technology that generates a partially ionized gas containing reactive oxygen and nitrogen species (RONS) at temperatures compatible with living tissue. CAP has demonstrated antimicrobial activity against a broad range of bacteria, including multidrug-resistant organisms, and has shown promise for wound decontamination and healing promotion.
When combined with LL-37, CAP therapy shows enhanced antimicrobial and wound healing effects. The RONS generated by CAP treatment can upregulate stress-response pathways in epithelial cells, including pathways that converge on CAMP gene expression. Additionally, the sub-lethal oxidative stress induced by CAP may increase bacterial membrane fluidity, enhancing LL-37's ability to insert into and disrupt bacterial membranes.
This triple-action approach - direct CAP-mediated bacterial killing, LL-37-mediated membrane disruption, and CAP-stimulated endogenous LL-37 production - represents an innovative multi-modal strategy for chronic wound management. Early-stage clinical studies of CAP therapy for wound healing have shown promising results, and the addition of LL-37 to these protocols could further improve outcomes.
Advanced Nanoparticle Delivery Systems
The development of nanoparticle delivery systems for LL-37 has progressed rapidly in recent years, with multiple platform technologies showing promise for improving the peptide's stability, targeting, and therapeutic efficacy.
Gold nanoparticles conjugated with LL-37 show enhanced antimicrobial activity compared to the free peptide, attributed to the increased local concentration of peptide at the nanoparticle-bacteria interface and the potential for gold-mediated photothermal effects when exposed to near-infrared light. Silver nanoparticles co-loaded with LL-37 provide dual antimicrobial mechanisms - the intrinsic antimicrobial activity of silver ions combined with LL-37's membrane-disrupting effects - creating a formulation that is effective at lower concentrations of both agents.
Polymeric nanoparticles based on PLGA (poly lactic-co-glycolic acid) or chitosan offer biodegradable, sustained-release delivery of LL-37 for wound healing applications. These nanoparticles protect the encapsulated peptide from proteolytic degradation, provide controlled release over days to weeks, and can be incorporated into wound dressings, hydrogels, or injectable formulations depending on the clinical application.
Lipid-based nanoparticles, including liposomes and solid lipid nanoparticles, mimic the membrane-like environment that stabilizes LL-37's alpha-helical structure, potentially preserving its antimicrobial activity during storage and delivery. Liposomal LL-37 formulations have shown enhanced stability at physiological temperature compared to the free peptide and improved penetration into biofilm matrices.
The most advanced nanoparticle delivery systems combine multiple features: protection from degradation, sustained release, targeted delivery (using surface ligands that bind to infected tissue markers), and stimuli-responsive release (triggered by pH, temperature, or enzymatic activity at the infection site). These "smart" delivery systems represent the cutting edge of LL-37 formulation technology and could eventually enable therapeutic applications that are not feasible with the unformulated peptide.
References
References
- Zanetti M. Cathelicidins, multifunctional peptides of the innate immunity. J Leukoc Biol. 2004;75(1):39-48. doi:10.1189/jlb.0403147
- Sorensen OE, Follin P, Johnsen AH, et al. Human cathelicidin, hCAP-18, is processed to the antimicrobial peptide LL-37 by extracellular cleavage with proteinase 3. Blood. 2001;97(12):3951-3959. doi:10.1182/blood.V97.12.3951
- Durr UH, Sudheendra US, Ramamoorthy A. LL-37, the only human member of the cathelicidin family of antimicrobial peptides. Biochim Biophys Acta. 2006;1758(9):1408-1425. doi:10.1016/j.bbamem.2006.03.030
- Wang G. Structures of human host defense cathelicidin LL-37 and its smallest antimicrobial peptide KR-12 in lipid micelles. J Biol Chem. 2008;283(47):32637-32643. doi:10.1074/jbc.M805533200
- Porcelli F, Verardi R, Shi L, Henzler-Wildman KA, Ramamoorthy A, Bhatt G. NMR structure of the cathelicidin-derived human antimicrobial peptide LL-37 in dodecylphosphocholine micelles. Biochemistry. 2008;47(20):5565-5572. doi:10.1021/bi702036s
- Sancho-Vaello E, Francois P, Bonber EJ, et al. The structure of the antimicrobial human cathelicidin LL-37 shows oligomerization and channel formation in the presence of membrane mimics. Sci Rep. 2020;10(1):17356. doi:10.1038/s41598-020-74401-5
- Xhindoli D, Pacor S, Benincasa M, Scocchi M, Gennaro R, Tossi A. The human cathelicidin LL-37 - A pore-forming antibacterial peptide and host-cell modulator. Biochim Biophys Acta. 2016;1858(3):546-566. doi:10.1016/j.bbamem.2015.11.003
- Wang TT, Nestel FP, Bourdeau V, et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J Immunol. 2004;173(5):2909-2912. doi:10.4049/jimmunol.173.5.2909
- Gombart AF, Borregaard N, Koeffler HP. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005;19(9):1067-1077. doi:10.1096/fj.04-3284com
- Dixon BM, Barker T, McKinnon T, et al. Positive correlation between circulating cathelicidin antimicrobial peptide (hCAP18/LL-37) and 25-hydroxyvitamin D levels in healthy adults. BMC Res Notes. 2012;5:575. doi:10.1186/1756-0500-5-575
- Mandic Havelka A, Yektaei-Karin E, Engblom L, et al. Vitamin D3 induces expression of human cathelicidin antimicrobial peptide 18 in newborns. Neonatology. 2010;97(2):182. doi:10.1159/000253816
- Schauber J, Oda Y, Buchau AS, et al. Histone acetylation in keratinocytes enables control of the expression of cathelicidin and CD14 by 1,25-dihydroxyvitamin D3. J Invest Dermatol. 2008;128(4):816-824. doi:10.1038/sj.jid.5701102
- Gronberg A, Mahlapuu M, Stahle M, Whately-Smith C, Rollman O. Treatment with LL-37 is safe and effective in enhancing healing of hard-to-heal venous leg ulcers: a randomized, placebo-controlled clinical trial. Wound Repair Regen. 2014;22(5):613-621. doi:10.1111/wrr.12211
- Heilborn JD, Nilsson MF, Kratz G, et al. The cathelicidin anti-microbial peptide LL-37 is involved in re-epithelialization of human skin wounds and is lacking in chronic ulcer epithelium. J Invest Dermatol. 2003;120(3):379-389. doi:10.1046/j.1523-1747.2003.12069.x
- de Breij A, Riool M, Cordfunke RA, et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci Transl Med. 2018;10(423):eaan4044. doi:10.1126/scitranslmed.aan4044
- Bowdish DM, Davidson DJ, Speert DP, Hancock RE. The human cationic peptide LL-37 induces activation of the extracellular signal-regulated kinase and p38 kinase pathways in primary human monocytes. J Immunol. 2004;172(6):3758-3765. doi:10.4049/jimmunol.172.6.3758
- Zheng Y, Niyonsaba F, Ushio H, et al. Cathelicidin LL-37 induces the generation of reactive oxygen species and release of human alpha-defensins from neutrophils. Br J Dermatol. 2007;157(6):1124-1131. doi:10.1111/j.1365-2133.2007.08196.x
- Lande R, Gregorio J, Facchinetti V, et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature. 2007;449(7162):564-569. doi:10.1038/nature06116
- Barlow PG, Svoboda P, Mackellar A, et al. Antiviral activity and increased host defense against influenza infection elicited by the human cathelicidin LL-37. PLoS One. 2011;6(10):e25333. doi:10.1371/journal.pone.0025333
- Tripathi S, Tecle T, Verma A, Crouch E, White M, Hartshorn KL. The human cathelicidin LL-37 inhibits influenza A viruses through a mechanism distinct from that of surfactant protein D or defensins. J Gen Virol. 2013;94(Pt 1):40-49. doi:10.1099/vir.0.045013-0
- Currie SM, Findlay EG, McHugh BJ, et al. The human cathelicidin LL-37 has antiviral activity against respiratory syncytial virus. PLoS One. 2013;8(8):e73659. doi:10.1371/journal.pone.0073659
- Overhage J, Campisano A, Bains M, Torfs EC, Rehm BH, Hancock RE. Human host defense peptide LL-37 prevents bacterial biofilm formation. Infect Immun. 2008;76(9):4176-4182. doi:10.1128/IAI.00318-08
- Noore J, Noore A, Li B. Cationic antimicrobial peptide LL-37 is effective against both extra- and intracellular Staphylococcus aureus. Antimicrob Agents Chemother. 2013;57(3):1283-1290. doi:10.1128/AAC.01650-12
- Dosler S, Karaaslan E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides. 2014;62:32-37. doi:10.1016/j.peptides.2014.09.021
- Yamasaki K, Di Nardo A, Bardan A, et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat Med. 2007;13(8):975-980. doi:10.1038/nm1616
- Kahlenberg JM, Kaplan MJ. Little peptide, big effects: the role of LL-37 in inflammation and autoimmune disease. J Immunol. 2013;191(10):4895-4901. doi:10.4049/jimmunol.1302005
- Schauber J, Gallo RL. Antimicrobial peptides and the skin immune defense system. J Allergy Clin Immunol. 2008;122(2):261-266. doi:10.1016/j.jaci.2008.03.027
- Vandamme D, Lanber B, Vergote I, et al. A comprehensive summary of LL-37, the factotum human cathelicidin peptide. Cell Immunol. 2012;280(1):22-35. doi:10.1016/j.cellimm.2012.11.009
- Ren SX, Shen J, Cheng AS, et al. The human cathelicidin peptide LL-37 inhibits pancreatic cancer growth by suppressing autophagy and reprogramming of the tumor immune microenvironment. Front Pharmacol. 2022;13:906625. doi:10.3389/fphar.2022.906625
- Bucki R, Leszczynska K, Namiot A, Sokolowski W. Cathelicidin LL-37: a multitask antimicrobial peptide. Arch Immunol Ther Exp. 2010;58(1):15-25. doi:10.1007/s00005-009-0057-2
- Nijnik A, Hancock RE. The roles of cathelicidin LL-37 in immune defences and novel clinical applications. Curr Opin Hematol. 2009;16(1):41-47. doi:10.1097/MOH.0b013e32831ac517
- Hancock RE, Haney EF, Gill EE. The immunology of host defence peptides: beyond antimicrobial activity. Nat Rev Immunol. 2016;16(5):321-334. doi:10.1038/nri.2016.29
- Fabisiak A, Murawska N, Fichna J. LL-37: Cathelicidin-related antimicrobial peptide with pleiotropic activity. Pharmacol Rep. 2016;68(4):802-808. doi:10.1016/j.pharep.2016.03.015
- Schauber J, Rieger D, Weiler F, et al. Heterogeneous expression of human cathelicidin hCAP18/LL-37 in inflammatory bowel diseases. Eur J Gastroenterol Hepatol. 2006;18(6):615-621. doi:10.1097/00042737-200606000-00007
- Bandurska K, Berdowska A, Barczynska-Felusiak R, Krupa P. Unique features of human cathelicidin LL-37. Biofactors. 2015;41(5):289-300. doi:10.1002/biof.1225
- Nagaoka I, Tamura H, Reich J. Therapeutic potential of cathelicidin peptide LL-37, an antimicrobial agent, in a murine sepsis model. Int J Mol Sci. 2020;21(17):5973. doi:10.3390/ijms21175973
- Chen X, Zou X, Qi G, et al. Roles and mechanisms of human cathelicidin LL-37 in cancer. Cell Physiol Biochem. 2018;47(3):1060-1073. doi:10.1159/000490183
- Mookherjee N, Anderson MA, Haagsman HP, Davidson DJ. Antimicrobial host defence peptides: functions and clinical potential. Nat Rev Drug Discov. 2020;19(5):311-332. doi:10.1038/s41573-019-0058-8
- Agier J, Efenberger M, Brzezinska-Blaszczyk E. Cathelicidin impact on inflammatory cells. Cent Eur J Immunol. 2015;40(2):225-235. doi:10.5114/ceji.2015.51359
- Ridyard KE, Overhage J. The potential of human peptide LL-37 as an antimicrobial and anti-biofilm agent. Antibiotics (Basel). 2021;10(6):650. doi:10.3390/antibiotics10060650
- Niyonsaba F, Kiatsurayanon C, Chieosilapatham P, Ogawa H. Friends or foes? Host defense (antimicrobial) peptides and proteins in human skin diseases. Exp Dermatol. 2017;26(11):989-998. doi:10.1111/exd.13314
