Executive Summary

Key Takeaways
- Figure 1: 5-Amino-1MQ targets the NNMT enzyme to shift metabolic balance away from fat storage and toward energy expenditure
- 5-Amino-1MQ (5-amino-1-methylquinolinium) is a small molecule inhibitor of nicotinamide N-methyltransferase (NNMT) that has emerged as a compelling research compound in the fight against obesity and metabolic dysfunction.
- Unlike traditional weight loss therapies that suppress appetite or block nutrient absorption, 5-Amino-1MQ works at the cellular level to reprogram how fat cells store and burn energy.
- The obesity epidemic continues to affect hundreds of millions of people worldwide, and current pharmacological options, while increasingly effective, still leave room for therapies that target different pathways.
- Semaglutide and tirzepatide have reshaped the weight loss landscape by acting on GLP-1 and GIP receptors in the brain and gut.
5-Amino-1MQ (5-amino-1-methylquinolinium) is a small molecule inhibitor of nicotinamide N-methyltransferase (NNMT) that has emerged as a compelling research compound in the fight against obesity and metabolic dysfunction. Unlike traditional weight loss therapies that suppress appetite or block nutrient absorption, 5-Amino-1MQ works at the cellular level to reprogram how fat cells store and burn energy.
The obesity epidemic continues to affect hundreds of millions of people worldwide, and current pharmacological options, while increasingly effective, still leave room for therapies that target different pathways. Semaglutide and tirzepatide have reshaped the weight loss landscape by acting on GLP-1 and GIP receptors in the brain and gut. But what if a compound could reduce fat mass without affecting appetite at all? That is precisely the promise of 5-Amino-1MQ.
NNMT is an enzyme found primarily in white adipose tissue (fat) and the liver. Its expression rises sharply with obesity. The enzyme performs a simple but metabolically consequential reaction: it transfers a methyl group from S-adenosylmethionine (SAM) to nicotinamide (a form of vitamin B3), producing 1-methylnicotinamide (1-MNA). This reaction has two downstream effects that promote fat storage. First, it diverts nicotinamide away from the NAD+ salvage pathway, reducing cellular NAD+ levels and lowering energy expenditure. Second, it depletes SAM, a universal methyl donor that controls gene expression through epigenetic modifications. When NNMT activity is high, cells enter a metabolic state that favors lipid accumulation and resists fat burning.
5-Amino-1MQ blocks this enzyme with high selectivity. In preclinical studies published in Biochemistry (2017) and subsequent follow-up work, the compound demonstrated remarkable properties. It is membrane-permeable, meaning it can cross cell walls to reach intracellular NNMT. It does not inhibit other SAM-dependent methyltransferases or enzymes in the NAD+ salvage pathway, giving it a clean pharmacological profile. And in cell culture experiments, it reduced intracellular 1-MNA production, boosted NAD+ levels, and suppressed the formation of new fat cells (adipogenesis).
The animal data have been equally striking. In diet-induced obese mice fed a high-fat diet, treatment with 5-Amino-1MQ for 11 weeks reduced body fat percentage from approximately 42% to 28%, a relative reduction of about one-third. Total body weight dropped by roughly 5%, white adipose tissue mass fell by 35%, and individual fat cell (adipocyte) size shrank by over 30%. These changes occurred without any measurable difference in food intake, confirming that the compound works through metabolic reprogramming rather than appetite suppression.
Beyond fat loss, NNMT inhibition has shown promise for muscle function. A 2024 study published in Scientific Reports found that 5-Amino-1MQ improved grip strength in aged mice by approximately 60% when combined with exercise training, compared to about 25% improvement from either intervention alone. This finding opens potential applications for sarcopenia, the age-related loss of muscle mass and strength that affects millions of older adults.
The compound's influence on NAD+ biology connects it to broader trends in longevity and metabolic health research. By preserving nicotinamide for conversion into NAD+ through the salvage pathway, 5-Amino-1MQ effectively acts as an NAD+ booster, a function it shares with compounds like NAD+ supplementation but through an entirely different mechanism. The preserved SAM also enhances polyamine metabolism and histone methylation, creating a cascade of metabolic benefits that extend beyond simple fat reduction.
Several important caveats apply. All efficacy data for 5-Amino-1MQ comes from preclinical research: cell culture studies and mouse models. No human clinical trials have been completed or, as of early 2026, formally registered. The compound has not received FDA approval for any indication. Practitioners who offer it do so through compounding pharmacies, and dosing protocols are derived from extrapolation of animal data and clinical experience rather than randomized controlled trials. Oral bioavailability appears to be lower than subcutaneous administration, though capsule formulations are the most common delivery method in practice.
This report examines every aspect of 5-Amino-1MQ in detail: the biology of NNMT and its role in metabolic regulation, the specific mechanism by which 5-Amino-1MQ inhibits this enzyme, the full body of preclinical evidence for fat loss, the NAD+ salvage pathway effects, the emerging data on muscle and exercise performance, practical dosing considerations, and the safety profile as currently understood. For those exploring the full spectrum of metabolic optimization tools, the GLP-1 research hub and peptide research hub provide additional context on complementary compounds.
NNMT Enzyme & Metabolic Regulation
Figure 2: NNMT enzyme activity in white adipose tissue and its central role in metabolic regulation
What Is NNMT?
Nicotinamide N-methyltransferase (NNMT) is a cytosolic enzyme that catalyzes one of the most metabolically significant methylation reactions in the body. It transfers a methyl group from S-adenosylmethionine (SAM) to nicotinamide (NAM), producing 1-methylnicotinamide (1-MNA) and S-adenosylhomocysteine (SAH) as byproducts. The enzyme was first characterized in the 1960s, but its role in metabolic disease did not become apparent until large-scale gene expression studies in the 2000s revealed that NNMT is dramatically upregulated in the white adipose tissue and liver of obese individuals and animal models of obesity.
The gene encoding NNMT sits on chromosome 11q23.1 in humans. Single nucleotide polymorphisms (SNPs) in the NNMT gene have been associated with body mass index, type 2 diabetes risk, hyperlipidemia, and hypertension in genome-wide association studies. These genetic correlations provided early evidence that NNMT was not merely a bystander in metabolic disease but a functional contributor. Expression levels in white adipose tissue correlate directly with percent body fat in both mice and humans, with obesity-prone strains showing high NNMT expression and obesity-resistant strains showing low expression (Kraus D, Yang Q, Kong D, et al. Nicotinamide N-methyltransferase knockdown protects against diet-induced obesity. Nature. 2014;508(7495):258-262. DOI: 10.1038/nature13198).
NNMT is expressed in virtually every tissue, but its metabolic effects are most pronounced in white adipose tissue, liver, and skeletal muscle. In lean individuals, NNMT activity in fat tissue is relatively low. As body fat accumulates and adipocytes enlarge, NNMT expression rises, creating a feed-forward loop: more NNMT leads to greater metabolic dysfunction, which promotes further fat accumulation, which drives even higher NNMT expression. Breaking this cycle is one of the key therapeutic rationales for NNMT inhibition.
The Dual Substrate Problem: NAM and SAM Depletion
Understanding why NNMT matters requires appreciating what happens to its two key metabolites. The enzyme consumes nicotinamide and SAM simultaneously, and the depletion of both molecules creates compounding metabolic problems.
Nicotinamide is the primary precursor for NAD+ through the salvage pathway, which produces approximately 85% of total cellular NAD+. In this pathway, the enzyme NAMPT converts nicotinamide into nicotinamide mononucleotide (NMN), which is then converted to NAD+ by NMNAT1-3 enzymes. NAD+ is essential for hundreds of metabolic reactions, including those that drive the citric acid cycle, fatty acid oxidation, and mitochondrial electron transport. When NNMT methylates nicotinamide to form 1-MNA, that nicotinamide can no longer be recycled into NAD+. The result is lower cellular NAD+ levels, reduced energy expenditure, and impaired fat oxidation.
SAM is the body's universal methyl donor, required for over 200 methylation reactions including DNA methylation, histone methylation, and the synthesis of creatine, phosphatidylcholine, and polyamines. When NNMT consumes SAM at elevated rates, the intracellular SAM/SAH ratio drops. This has direct consequences for gene expression: histone methylation patterns shift, and genes involved in energy expenditure and polyamine metabolism become less active. The polyamine connection is particularly important. SAM is required for the synthesis of decarboxylated SAM (dcSAM), which donates aminopropyl groups during the conversion of putrescine to spermidine and spermidine to spermine. These polyamine interconversion reactions, collectively called polyamine flux, consume acetyl-CoA and generate reactive oxygen species that drive energy expenditure. When NNMT depletes SAM, polyamine flux slows, and the metabolic rate in fat tissue drops.
NNMT in Adipose Tissue Biology
White adipose tissue is not a passive storage depot. It is an active endocrine organ that secretes hormones (adipokines), responds to insulin signaling, and dynamically adjusts its metabolic activity based on energy balance. NNMT plays a central role in determining whether adipose tissue is metabolically active or quiescent.
During adipogenesis, the process by which precursor cells differentiate into mature fat cells, NNMT is transactivated by CCAAT/Enhancer Binding Protein beta (C/EBP-beta) in response to glucocorticoid signaling (Esteves M, Maguire R, et al. Nicotinamide N-methyltransferase (NNMT) regulates the glucocorticoid signaling pathway during the early phase of adipogenesis. Scientific Reports. 2023;13(1):8049. DOI: 10.1038/s41598-023-34916-z). This positions NNMT as a key component of the glucocorticoid-C/EBP axis that drives the early stages of fat cell formation. Blocking NNMT at this stage can suppress adipogenesis itself, preventing new fat cells from forming.
In mature adipocytes, NNMT activity determines the metabolic phenotype of the cell. Adipocytes with high NNMT have lower NAD+ levels, reduced sirtuin (SIRT1) activity, and lower rates of fatty acid oxidation. They store more lipid and have larger lipid droplets. Adipocytes with low NNMT activity (either naturally or through pharmacological inhibition) have higher NAD+ levels, more active SIRT1 signaling, and burn more fat. They are metabolically healthier and smaller in size.
The 2014 Nature study by Kraus and colleagues provided the most compelling evidence for this relationship. Using antisense oligonucleotides to knock down NNMT expression in white adipose tissue and liver, they showed that treated mice on a high-fat diet had significantly less body fat, smaller adipocytes, higher energy expenditure, better glucose tolerance, and lower circulating lipid levels compared to controls. The treated mice were essentially protected from the metabolic consequences of a high-fat diet. Mechanistically, NNMT knockdown increased adipose SAM and NAD+ levels, activated polyamine flux (measured by increased urinary diacetylspermine excretion), and enhanced histone H3 lysine 4 (H3K4) methylation at gene promoters involved in energy expenditure.
NNMT in Liver and Metabolic Syndrome
While adipose tissue gets the most attention in obesity research, NNMT in the liver also plays a significant role. Hepatic NNMT expression rises with obesity and contributes to the development of non-alcoholic fatty liver disease (NAFLD) and insulin resistance. In the liver, NNMT activity reduces NAD+ availability for enzymes like SIRT1 and SIRT3 that are critical for mitochondrial function and fatty acid oxidation. This creates conditions favorable for hepatic lipid accumulation (steatosis).
The relationship between NNMT and hepatic steatosis is somewhat nuanced, however. A 2015 review in Cell Metabolism noted that while NNMT is detrimental in adipose tissue, its role in the liver may be more complex, with some studies suggesting that NNMT-generated 1-MNA has anti-inflammatory properties in certain hepatic contexts. This does not diminish the therapeutic rationale for NNMT inhibition in obesity, but it does suggest that the ideal approach might involve tissue-selective targeting. 5-Amino-1MQ, as a systemic inhibitor, blocks NNMT in all tissues, and the net metabolic effect in preclinical studies has been overwhelmingly positive.
NNMT also intersects with the broader metabolic syndrome through its effects on insulin signaling. In adipocytes, reduced NAD+ impairs insulin-stimulated glucose uptake by decreasing SIRT1-mediated deacetylation of insulin receptor substrate proteins. In the liver, reduced NAD+ impairs gluconeogenic regulation. These effects contribute to the insulin resistance and hyperglycemia that characterize type 2 diabetes. Genetic studies have confirmed these links: single nucleotide variants in the NNMT gene are significantly associated with disturbances in energy metabolism, obesity, type 2 diabetes, hyperlipidemia, and hypertension (Liu Y, Liang J, et al. Roles of nicotinamide N-methyltransferase in obesity and type 2 diabetes. BioMed Research International. 2021;2021:9924314. DOI: 10.1155/2021/9924314).
NNMT as a Metabolic Crossroads
What makes NNMT such an attractive therapeutic target is its position at the intersection of multiple metabolic pathways. It simultaneously affects NAD+ metabolism, the methionine cycle, polyamine flux, and epigenetic regulation. Inhibiting a single enzyme produces effects across all of these domains. This is unusual in pharmacology, where most drugs target a single pathway and have limited upstream consequences.
A 2024 review in Frontiers in Pharmacology described NNMT as standing at the crossroads between cellular metabolism and epigenetic regulation (Wang Y, Zhou X, et al. Nicotinamide N-methyltransferase (NNMT): a novel therapeutic target for metabolic syndrome. Frontiers in Pharmacology. 2024;15:1410479. DOI: 10.3389/fphar.2024.1410479). The review catalogued NNMT's involvement in obesity, type 2 diabetes, cardiovascular disease, and even cancer, where high NNMT expression in tumors is associated with poor prognosis and resistance to therapy. While the cancer applications are beyond the scope of this report, they illustrate the breadth of NNMT's biological influence and reinforce its importance as a drug target.
For those interested in related metabolic targets, compounds like AOD-9604 and Fragment 176-191 also target fat metabolism through different mechanisms, while tesofensine approaches weight loss through neurotransmitter reuptake inhibition. The drug comparison hub covers how these approaches differ in their metabolic targets and clinical evidence.
NNMT Expression Across the Lifespan
NNMT activity is not static. It changes with age, hormonal status, and metabolic state. In aging, NNMT expression in skeletal muscle increases, contributing to the decline in NAD+ that has been linked to age-related muscle wasting (sarcopenia), mitochondrial dysfunction, and reduced exercise capacity. This age-related increase in NNMT provides a mechanistic explanation for why older adults have lower NAD+ levels and less metabolic flexibility, even when controlling for body composition.
Glucocorticoid levels, which rise with chronic stress and certain medical conditions (Cushing's syndrome, long-term corticosteroid use), also increase NNMT expression. This creates a direct link between stress hormones and fat accumulation, particularly visceral fat, which expresses NNMT at higher levels than subcutaneous fat. The cortisol-NNMT-adipogenesis axis helps explain why chronic stress is so strongly associated with central obesity and metabolic syndrome.
Sex differences in NNMT expression have also been observed, though the data are less consistent. Some studies report higher NNMT activity in male adipose tissue, which could partially explain sex-based differences in fat distribution and metabolic disease risk. More research is needed in this area to draw definitive conclusions.
Mechanism of Action
Chemical Structure and Pharmacological Properties
5-Amino-1MQ has the chemical formula C10H11N2+ and a molecular weight of approximately 159.2 g/mol. It belongs to the quinolinium class of compounds, characterized by a positively charged nitrogen in the aromatic ring system. The "5-amino" designation refers to an amino group at the 5-position of the quinoline ring, while the "1-methyl" refers to a methyl group on the ring nitrogen. This specific substitution pattern was identified through structure-activity relationship studies as conferring both high NNMT inhibitory potency and excellent membrane permeability.
The compound was first described in detail by Neelakantan H, Wang HY, Vance V, et al. in their 2017 publication in Biochemistry (Neelakantan H, Wang HY, Vance V, et al. Selective and membrane-permeable small molecule inhibitors of nicotinamide N-methyltransferase reverse high fat diet-induced obesity in mice. Biochemistry. 2018;57(5):481-486. DOI: 10.1021/acs.biochem.7b01107). The researchers screened a series of nicotinamide analogs for NNMT inhibitory activity and identified 5-Amino-1MQ as the most potent and selective compound in their series. Critically, 5-Amino-1MQ demonstrated high membrane permeability in bidirectional Caco-2 cell permeation assays, a standard test for predicting oral absorption and cellular uptake.
The selectivity profile is a key strength. 5-Amino-1MQ does not inhibit other SAM-dependent methyltransferases, which is important because there are hundreds of methyltransferases in human cells that perform essential functions in DNA methylation, histone modification, and small molecule metabolism. Non-selective methyltransferase inhibition would produce widespread and unpredictable effects. Similarly, 5-Amino-1MQ does not inhibit enzymes in the NAD+ salvage pathway itself (NAMPT, NMNAT1-3), meaning it preserves nicotinamide for NAD+ synthesis without interfering with the enzymatic machinery that performs the conversion.
Competitive Inhibition of NNMT
5-Amino-1MQ works as a competitive inhibitor at the nicotinamide binding site of NNMT. The enzyme normally binds nicotinamide and SAM simultaneously, bringing them into close proximity for the methyl transfer reaction. 5-Amino-1MQ occupies the nicotinamide binding pocket, preventing the natural substrate from accessing the active site. Because the compound has a higher affinity for the binding site than nicotinamide itself, it effectively displaces the natural substrate even when nicotinamide concentrations are within normal physiological range.
The kinetics of this inhibition are concentration-dependent. At low concentrations, some NNMT molecules remain uninhibited and continue to methylate nicotinamide. As the concentration of 5-Amino-1MQ increases, a greater fraction of NNMT molecules are occupied by the inhibitor, and the rate of 1-MNA production falls proportionally. Complete inhibition of all NNMT activity is neither achievable nor desirable at physiological doses; rather, the goal is to reduce NNMT activity enough to shift the metabolic balance toward greater NAD+ production and SAM preservation.
The downstream consequences of this inhibition unfold across several interconnected pathways.
Pathway 1: NAD+ Restoration
When NNMT is inhibited, nicotinamide that would have been methylated to 1-MNA instead remains available for the NAD+ salvage pathway. NAMPT converts this nicotinamide to NMN, and NMN is then converted to NAD+ by NMNAT enzymes. The result is higher intracellular NAD+ concentrations.
Elevated NAD+ activates a family of enzymes called sirtuins, particularly SIRT1 and SIRT3. SIRT1 is a NAD+-dependent deacetylase that activates PGC-1alpha (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), the master regulator of mitochondrial biogenesis. PGC-1alpha activation increases the number and activity of mitochondria, enhances fatty acid oxidation, and promotes the conversion of white adipose tissue toward a more metabolically active phenotype. SIRT3, located in the mitochondrial matrix, deacetylates and activates multiple enzymes in the electron transport chain and fatty acid oxidation pathway, directly increasing cellular energy production.
This NAD+ restoration mechanism is shared conceptually with other NAD+ boosting strategies, including NMN supplementation, NR (nicotinamide riboside) supplementation, and direct NAD+ administration. However, 5-Amino-1MQ achieves NAD+ elevation through a fundamentally different approach: rather than providing more precursor, it prevents the loss of precursor that already exists. This "conservation" strategy may prove more efficient in tissues where NNMT activity is high, because the locally generated nicotinamide is preserved exactly where it is needed.
Pathway 2: SAM Preservation and Epigenetic Effects
The second major consequence of NNMT inhibition is the preservation of SAM, the universal methyl donor. With less SAM being consumed by NNMT, the intracellular SAM/SAH ratio increases. This ratio is the primary determinant of methylation capacity within the cell.
Higher SAM availability has direct effects on histone methylation. In the 2014 Kraus Nature study, NNMT knockdown in adipose tissue increased trimethylation of histone H3 at lysine 4 (H3K4me3) at the promoters of genes involved in polyamine metabolism. H3K4me3 is an activating mark, meaning that its presence turns on gene expression. The genes activated by this epigenetic change include ornithine decarboxylase (ODC) and spermidine/spermine N1-acetyltransferase (SSAT), both of which are key enzymes in the polyamine flux pathway.
This epigenetic reprogramming is not a transient effect. Once histone methylation patterns are established, they tend to persist through cell divisions, creating lasting changes in gene expression that outlive the immediate presence of the inhibitor. This raises the possibility that a course of NNMT inhibition could produce metabolic benefits that persist even after treatment ends, though this hypothesis has not been formally tested in chronic dosing studies.
Pathway 3: Polyamine Flux Activation
The polyamine pathway is the most novel aspect of NNMT inhibition and the one that most directly drives the increase in energy expenditure observed in preclinical studies. Polyamines (putrescine, spermidine, and spermine) are small organic cations found in all living cells. Their metabolism involves a series of interconversion reactions that collectively consume ATP, acetyl-CoA, and oxygen while generating reactive oxygen species and CO2.
Here is how it works. SAM is decarboxylated by SAM decarboxylase to form dcSAM. ODC converts ornithine to putrescine. dcSAM donates an aminopropyl group to putrescine (via spermidine synthase) to form spermidine, and another aminopropyl group to spermidine (via spermine synthase) to form spermine. SSAT acetylates spermine and spermidine, and polyamine oxidase (PAO) then oxidizes the acetylated products back to lower-order polyamines. This cycle, the polyamine flux, is an energy-consuming process that generates heat.
When NNMT is inhibited and SAM levels rise, more dcSAM is produced, and the polyamine flux accelerates. The Kraus study showed that NNMT knockdown increased ODC and SSAT activity, increased the expression of both enzymes at the mRNA level, and increased urinary excretion of diacetylspermine (a polyamine metabolite that serves as a biomarker of flux activity). Oxygen consumption in adipocytes increased in an ODC-, SSAT-, and PAO-dependent manner, confirming that the polyamine flux was the direct driver of enhanced energy expenditure.
This mechanism is distinct from any currently approved weight loss therapy. GLP-1 agonists like semaglutide reduce appetite and slow gastric emptying. Tirzepatide adds GIP receptor activation. Retatrutide adds glucagon receptor activation. But none of these agents directly increase adipose energy expenditure through polyamine metabolism. This makes 5-Amino-1MQ a mechanistically unique compound with potential for combination therapy.
Pathway 4: Suppression of Adipogenesis
Beyond affecting existing fat cells, 5-Amino-1MQ suppresses the formation of new fat cells. In cell culture studies, treatment with the compound reduced the differentiation of preadipocytes (precursor cells) into mature adipocytes. This anti-adipogenic effect appears to be mediated by the disruption of the glucocorticoid-C/EBP-beta-NNMT axis that normally drives early adipocyte differentiation.
By blocking NNMT during the critical early phase of adipogenesis, 5-Amino-1MQ prevents the full activation of the adipogenic transcription factor cascade that includes C/EBP-beta, C/EBP-alpha, and PPAR-gamma. Without the complete activation of this cascade, preadipocytes fail to differentiate into lipid-laden mature fat cells. This effect complements the metabolic reprogramming of existing adipocytes: fewer new fat cells are created while existing fat cells become smaller and more metabolically active.
Pathway 5: Reduced Lipogenesis
In addition to suppressing the creation of new fat cells, 5-Amino-1MQ directly reduces lipogenesis (the synthesis of new fat) within existing adipocytes. In vitro studies showed that NNMT inhibition decreased the expression of lipogenic enzymes including fatty acid synthase (FASN) and acetyl-CoA carboxylase (ACC). These enzymes catalyze the conversion of excess carbohydrates into fatty acids for storage. With their expression reduced, adipocytes shift from a net fat-storing phenotype to a net fat-burning phenotype.
The combined effect of all five pathways creates a comprehensive anti-obesity mechanism: higher NAD+ activates fat-burning enzymes, preserved SAM drives epigenetic changes that increase energy expenditure through polyamine flux, adipogenesis is suppressed, and lipogenesis is reduced. The net result, observed consistently in preclinical studies, is smaller fat cells, less total body fat, and higher metabolic rate, all without changes in food intake.
Pharmacokinetic Considerations
As a small, positively charged molecule, 5-Amino-1MQ has pharmacokinetic properties that differ from peptide-based therapies. Its membrane permeability was confirmed in Caco-2 cell assays, which model intestinal absorption. This suggests reasonable oral bioavailability, though formal pharmacokinetic studies with plasma concentration curves have not been published in the peer-reviewed literature.
The compound's small size (MW ~159 Da) gives it advantages in tissue distribution. Unlike large peptides or proteins, it can readily cross cell membranes to reach its intracellular target. NNMT is a cytosolic enzyme, not a cell surface receptor, so any effective inhibitor must be able to enter cells. This requirement for membrane permeability was a key design criterion in the development of 5-Amino-1MQ and distinguishes it from earlier NNMT inhibitors that were potent in enzyme assays but unable to cross cell membranes.
The half-life, volume of distribution, and metabolic clearance pathways for 5-Amino-1MQ have not been fully characterized in published studies. Practitioners generally recommend daily dosing, suggesting that the effective duration of action is less than 24 hours. The compound appears to be cleared relatively quickly, as NNMT inhibition needs to be maintained through repeated dosing rather than producing long-lasting enzyme inactivation.
Preclinical Fat Loss Data
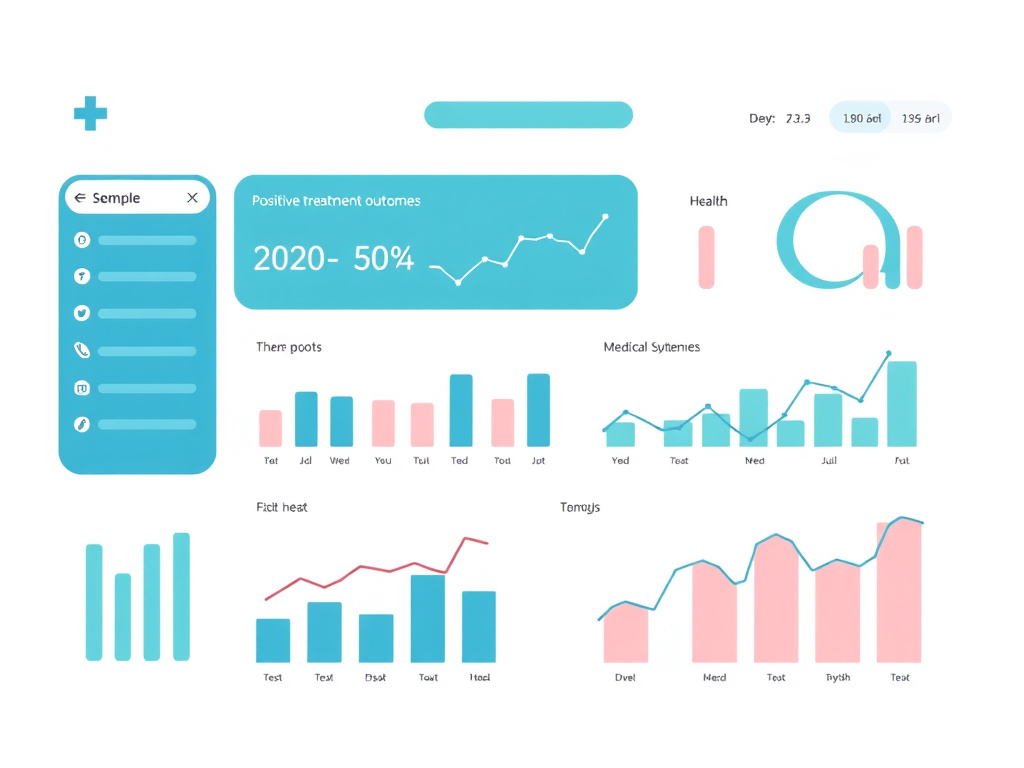
Body Fat % After 11 Weeks (High-Fat Diet Model)
The Foundational Diet-Induced Obesity Study
The primary preclinical evidence for 5-Amino-1MQ's fat loss effects comes from the work of Neelakantan and Wahlberg published in Biochemistry (2018), which built on the foundational NNMT knockdown studies by Kraus et al. in Nature (2014). The 2018 study was the first to test a pharmacological NNMT inhibitor, as opposed to genetic knockdown, in a diet-induced obesity (DIO) mouse model.
In this study, male C57BL/6 mice were placed on a high-fat diet (60% calories from fat) for an extended period to induce obesity, a standard model that reliably produces the metabolic phenotype seen in human obesity: excess body fat, enlarged adipocytes, insulin resistance, and dyslipidemia. After obesity was established, mice were randomized to receive either 5-Amino-1MQ or vehicle control while continuing on the high-fat diet.
The treatment lasted 11 weeks. During this period, the researchers measured body weight weekly, assessed food intake, and at the end of the study collected adipose tissue for histological analysis (adipocyte sizing), weighed fat depots, and measured body composition. The results were striking across every endpoint measured.
Body Weight and Fat Mass Results
Mice treated with 5-Amino-1MQ showed a 5% reduction in total body weight compared to vehicle-treated controls despite continuing on the same high-fat diet. While a 5% reduction in total body weight may sound modest, the composition of that weight loss was heavily skewed toward fat tissue. White adipose tissue mass decreased by approximately 35%, indicating that the compound was specifically targeting fat stores rather than causing generalized weight loss from muscle or organ tissue.
Body fat percentage in the control group (high-fat diet, no treatment) reached approximately 42% by the end of the 11-week period. In the 5-Amino-1MQ treated group, body fat percentage was approximately 28%, representing a relative reduction of about one-third. This magnitude of fat reduction, achieved without caloric restriction or increased physical activity, was remarkable for a single-agent pharmacological intervention.
Individual adipocyte size shrank by more than 30% in treated mice, and adipocyte volume decreased by approximately 40%. These histological findings confirmed that the fat loss was not due to adipocyte death (which would cause inflammation and fibrosis) but rather to metabolic shrinkage of existing fat cells. The adipocytes simply stored less lipid because they were burning more energy through the polyamine flux and NAD+-dependent pathways described in the mechanism section.
Food Intake Was Unchanged
One of the most significant findings was that food intake did not differ between treated and control groups. This observation is critical for several reasons. First, it confirms that 5-Amino-1MQ works through metabolic reprogramming rather than appetite suppression. The mice ate the same amount of high-fat food but stored less of it as body fat. Second, it differentiates the compound mechanistically from GLP-1 receptor agonists like semaglutide and liraglutide, which achieve much of their weight loss effect through reduced food intake. Third, it suggests that 5-Amino-1MQ could be combined with appetite-suppressing agents for additive or complementary effects, since the two approaches would work through independent pathways.
The lack of appetite effects also speaks to the compound's selectivity. Many metabolic interventions that increase energy expenditure (thyroid hormone, DNP, high-dose caffeine) simultaneously activate compensatory hunger signals through hypothalamic circuits. The absence of compensatory hyperphagia with 5-Amino-1MQ suggests that the increased energy expenditure it produces does not trigger the hypothalamic energy-sensing mechanisms that normally defend body weight against depletion.
Metabolic Markers
Beyond body composition changes, treated mice showed improvements in several metabolic markers. Intracellular NAD+ levels in adipose tissue were significantly elevated, confirming the mechanistic prediction that NNMT inhibition would preserve nicotinamide for NAD+ synthesis. 1-Methylnicotinamide (1-MNA) levels in adipose tissue were correspondingly reduced, confirming that NNMT enzymatic activity was effectively inhibited by the compound.
Lipogenesis was suppressed in treated adipocytes, with reduced expression of key lipogenic enzymes. This metabolic shift from lipid synthesis to lipid oxidation is consistent with the increased NAD+ and SAM levels driving sirtuin activation and polyamine flux, respectively.
The Combined Diet Switch and NNMT Inhibition Study
A 2022 follow-up study published in Scientific Reports examined what happens when NNMT inhibition is combined with a dietary intervention. In this experiment, diet-induced obese mice were either maintained on the high-fat diet, switched to a low-fat diet (diet switch alone), treated with 5-Amino-1MQ while remaining on the high-fat diet (treatment alone), or given the combination of diet switch plus 5-Amino-1MQ treatment (Kannt A, Rajagopal S, et al. Reduced calorie diet combined with NNMT inhibition establishes a distinct microbiome in DIO mice. Scientific Reports. 2022;12(1):843. DOI: 10.1038/s41598-021-03670-5).
The combination group showed the most dramatic results. Body weight and whole-body adiposity normalized rapidly to levels comparable to age-matched lean control mice that had never been on a high-fat diet. This complete normalization was achieved faster and more completely than with diet switch alone, which, even after the same time period, was unable to fully restore body composition to lean control levels.
This finding carries important translational implications. In human obesity treatment, dietary changes alone often fail to produce complete normalization of body composition, partly because expanded adipose tissue resists shrinkage through metabolic adaptations that lower energy expenditure (the so-called "metabolic adaptation" or "starvation response"). By simultaneously increasing adipose energy expenditure through NNMT inhibition, 5-Amino-1MQ may overcome this adaptation and allow dietary changes to produce their full potential effect.
Microbiome Effects
The same 2022 study also examined changes in the gut microbiome, an increasingly recognized player in metabolic health. Both diet switch and NNMT inhibition independently altered the gut microbial community, but the combination of both interventions produced a distinct microbiome signature that was different from either intervention alone.
Specifically, the combination treatment increased the abundance of bacterial genera associated with lean metabolic phenotypes and reduced the abundance of genera associated with obesity and metabolic dysfunction. While the causal direction of these changes (whether the altered microbiome contributes to the metabolic improvement or is simply a consequence of it) remains unclear, the findings suggest that NNMT inhibition may have beneficial effects on the gut-metabolic axis that extend beyond direct adipose tissue mechanisms.
This is an area where larazotide, a compound that modulates gut permeability, and other gut-targeted therapies may eventually intersect with NNMT inhibition research. The gut microbiome's role in obesity and metabolic disease is an active area of investigation across multiple therapeutic modalities.
Comparison to NNMT Genetic Knockdown Results
How does pharmacological NNMT inhibition with 5-Amino-1MQ compare to genetic NNMT knockdown? The 2014 Kraus Nature study used antisense oligonucleotides (ASOs) to reduce NNMT mRNA levels by approximately 50-80% in adipose tissue and liver. The metabolic effects were substantial: protection from diet-induced obesity, increased energy expenditure, improved glucose tolerance, reduced circulating lipids, and reduced hepatic steatosis.
The pharmacological approach with 5-Amino-1MQ produces qualitatively similar results, though the magnitude of effect may differ depending on the degree of NNMT inhibition achieved at a given dose. ASO-mediated knockdown reduces NNMT protein levels over days to weeks and maintains suppression for the duration of treatment. Small molecule inhibition with 5-Amino-1MQ blocks enzyme activity immediately but requires sustained plasma concentrations to maintain the effect. Despite these pharmacological differences, both approaches converge on the same downstream outcomes: smaller fat cells, less total body fat, and improved metabolic markers.
One advantage of the pharmacological approach is reversibility. If adverse effects emerge, 5-Amino-1MQ can be discontinued and NNMT activity will return to baseline as the drug is cleared. Genetic knockdown approaches are less easily reversed. This reversibility is a practical advantage for any investigational compound, as it allows for dose titration and provides a safety margin.
Dose-Response Relationships
Detailed dose-response data for 5-Amino-1MQ in animal models have not been extensively published. The primary studies used a single dose level selected based on in vitro potency and preliminary pharmacokinetic estimates. Establishing a clear dose-response relationship (showing that higher doses produce greater effects up to a ceiling, and identifying the minimum effective dose) would be an important step toward clinical development.
What is known from in vitro data is that the degree of NNMT inhibition increases with drug concentration in a predictable, concentration-dependent manner. At the concentrations achieved in the animal studies, NNMT activity was substantially but not completely inhibited, suggesting that there may be room for greater efficacy at higher doses. Whether this would come at the cost of increased side effects is unknown.
Translational Considerations: Mice to Humans
Translating preclinical fat loss data from mice to humans requires caution. Mice have a much higher metabolic rate per unit body mass than humans (approximately 7-10 times higher), meaning that any intervention that increases energy expenditure will produce proportionally larger effects in mice. A 5% body weight reduction in mice on a high-fat diet might translate to a smaller percentage reduction in humans, or might require higher relative doses to achieve equivalent results.
Additionally, the high-fat diet model in mice produces a relatively extreme metabolic phenotype (60% calories from fat is far higher than most human diets). The response to NNMT inhibition in a more moderate metabolic environment may differ. Against these cautions, the consistency of the preclinical findings across multiple studies and endpoints, and the strong biological rationale connecting NNMT to multiple obesity-related pathways, provide reasonable confidence that some degree of translation to humans is likely.
For context, other weight loss compounds showed impressive preclinical results that translated to meaningful human efficacy. Semaglutide reduced body weight by approximately 20% in mice and produces 10-15% weight loss in humans. Tirzepatide showed similar preclinical-to-clinical translation. Whether 5-Amino-1MQ will follow this pattern remains to be determined through formal clinical trials.
What the Data Doesn't Show
Honest assessment of the preclinical data requires acknowledging its limitations. The studies were conducted in a single mouse strain (C57BL/6) using a single obesity model (high-fat diet). Results in other strains, in genetically obese models (ob/ob, db/db), or in female mice have not been published. The treatment duration was limited to 11 weeks; long-term efficacy and safety over months to years are unknown. Metabolic endpoints beyond body composition (cardiovascular function, hepatic function, renal function) were not comprehensively assessed. And the studies were conducted by a relatively small group of researchers; independent replication by unaffiliated laboratories would strengthen confidence in the findings.
These limitations do not invalidate the existing data but place it in the appropriate context of early-stage research. The compound is promising but unproven in humans, and the preclinical evidence, while consistent and mechanistically coherent, represents the starting point of a therapeutic development process rather than its completion. The science and research page provides broader context on the evidence standards applied to different compounds in the FormBlends research library.
NAD+ Salvage Pathway Effects
NAD+ Biology: The Cellular Energy Currency
Nicotinamide adenine dinucleotide (NAD+) is among the most important molecules in human biology. It participates as a coenzyme in over 500 enzymatic reactions, making it indispensable for energy metabolism, DNA repair, gene expression regulation, and cellular stress responses. NAD+ exists in two forms: the oxidized form (NAD+) and the reduced form (NADH). The NAD+/NADH ratio reflects the cell's redox state and drives the direction of metabolic reactions including glycolysis, the citric acid cycle, and mitochondrial oxidative phosphorylation.
NAD+ levels decline with age, obesity, and metabolic disease. This decline has been linked to virtually every hallmark of aging: mitochondrial dysfunction, genomic instability, cellular senescence, stem cell exhaustion, and altered intercellular communication. Restoring NAD+ levels has become one of the most actively pursued strategies in longevity research, with multiple approaches under investigation including NMN supplementation, NR (nicotinamide riboside) supplementation, CD38 inhibition, and NNMT inhibition.
The Three NAD+ Biosynthesis Pathways
Cells produce NAD+ through three distinct pathways. Understanding each is necessary to appreciate where NNMT inhibition fits in the broader NAD+ landscape.
The de novo biosynthesis pathway (also called the kynurenine pathway) converts tryptophan, an essential amino acid, into NAD+ through a series of eight enzymatic steps. This pathway is the only one that can generate NAD+ from scratch, but it is relatively slow and operates primarily in the liver. It contributes a small fraction of total NAD+ production under normal conditions.
The Preiss-Handler pathway converts nicotinic acid (another form of vitamin B3) into NAD+ in three steps. Nicotinic acid is converted to nicotinic acid mononucleotide by nicotinic acid phosphoribosyltransferase (NAPRT), then to nicotinic acid adenine dinucleotide (NaAD) by NMNAT enzymes, and finally to NAD+ by NAD+ synthetase. This pathway is active in tissues that express NAPRT and is the target of niacin (nicotinic acid) supplementation strategies.
The salvage pathway is by far the most important, producing approximately 85% of total cellular NAD+. In this pathway, nicotinamide (which is released when NAD+-consuming enzymes like sirtuins, PARPs, and CD38 use NAD+ as a substrate) is recycled back to NAD+ in two steps. First, NAMPT (nicotinamide phosphoribosyltransferase, the rate-limiting enzyme) converts nicotinamide to NMN. Then NMNAT enzymes convert NMN to NAD+. This recycling process is extremely efficient, allowing a relatively small pool of nicotinamide to support the high NAD+ turnover required by active cells.
NNMT as the NAD+ Salvage Pathway's Enemy
NNMT directly competes with the salvage pathway for nicotinamide. Every molecule of nicotinamide that NNMT methylates to form 1-MNA is a molecule that can no longer enter the salvage pathway to become NAD+. In tissues with high NNMT expression (obese adipose tissue, for example), this diversion can be substantial, meaningfully reducing the cellular NAD+ pool.
The competition between NNMT and NAMPT for nicotinamide is the critical regulatory point. NAMPT operates with a Km (Michaelis constant) for nicotinamide of approximately 2-5 micromolar, meaning it efficiently converts nicotinamide to NMN at low substrate concentrations. NNMT's Km for nicotinamide is somewhat higher, but its expression level in obese adipose tissue is so elevated that total enzyme activity can substantially deplete the local nicotinamide pool. The result is a tissue-specific NAD+ deficit that impairs mitochondrial function, reduces sirtuin activity, and promotes the metabolically sluggish adipocyte phenotype characteristic of obesity.
By inhibiting NNMT, 5-Amino-1MQ tilts this competition decisively in favor of the salvage pathway. More nicotinamide is available for NAMPT, more NMN is produced, and more NAD+ is generated. The in vitro data confirm this: treatment of adipocytes with 5-Amino-1MQ significantly increased intracellular NAD+ levels while reducing 1-MNA, demonstrating that the substrate was being redirected from the methylation pathway to the salvage pathway.
Downstream Effects of NAD+ Restoration in Adipose Tissue
The increase in adipose NAD+ produced by NNMT inhibition activates several downstream pathways that contribute to fat loss and metabolic improvement.
SIRT1 Activation: SIRT1 is a NAD+-dependent deacetylase that is among the most studied longevity-associated proteins. In adipose tissue, SIRT1 deacetylates and activates PGC-1alpha, promoting mitochondrial biogenesis and fatty acid oxidation. It also deacetylates PPAR-gamma, the master transcription factor for adipogenesis, shifting its activity from a pro-storage to a pro-mobilization phenotype. SIRT1 additionally deacetylates NF-kB, reducing inflammatory signaling in adipose tissue. Chronic low-grade inflammation in fat tissue (adipose tissue inflammation) is a hallmark of obesity and contributes to insulin resistance; SIRT1-mediated anti-inflammatory effects may help resolve this.
SIRT3 Activation: In mitochondria, SIRT3 uses NAD+ to deacetylate and activate multiple enzymes in the electron transport chain (complexes I, II, and III), the TCA cycle (isocitrate dehydrogenase 2, succinate dehydrogenase), and fatty acid oxidation (long-chain acyl-CoA dehydrogenase). The net effect is increased mitochondrial efficiency and greater fat oxidation capacity. This is particularly relevant in adipose tissue, where mitochondrial dysfunction is a feature of obesity and contributes to the metabolic inflexibility that makes weight loss difficult.
PARP-1 Function: PARPs (poly ADP-ribose polymerases) are NAD+-consuming enzymes involved in DNA repair. In obesity, DNA damage in adipose tissue is elevated due to oxidative stress, and PARP activation further depletes already-low NAD+ pools. By restoring NAD+ through NNMT inhibition, cells can better support both sirtuin signaling and DNA repair simultaneously, rather than facing a competition for limited NAD+ substrate.
Comparison with Other NAD+ Boosting Strategies
5-Amino-1MQ occupies a unique niche among NAD+ boosting strategies. Let's compare the major approaches.
| Strategy | Mechanism | Target Tissue | Clinical Evidence | Oral Availability |
|---|---|---|---|---|
| NMN Supplementation | Provides direct NAD+ precursor | Systemic | Several human trials completed | Yes (capsules) |
| NR (Nicotinamide Riboside) | Provides NAD+ precursor via NRK pathway | Systemic | Multiple human trials completed | Yes (capsules) |
| NAD+ IV/Injection | Direct NAD+ administration | Systemic | Limited human data | No (IV/injection) |
| NAD+ Nasal | Direct NAD+ via nasal mucosa | Systemic/CNS | Limited human data | Nasal spray |
| CD38 Inhibition | Reduces NAD+ consumption | Systemic | Preclinical only | Varies |
| 5-Amino-1MQ (NNMT Inhibition) | Preserves NAM for salvage pathway | Primarily adipose/liver | Preclinical only | Yes (capsules) |
The key distinction is that NMN and NR work by flooding the system with additional precursor, while 5-Amino-1MQ works by preventing the loss of precursor that already exists. In tissues with high NNMT expression (adipose tissue in obese individuals), the conservation approach may be more efficient than the supplementation approach because it targets the specific metabolic bottleneck in that tissue. In tissues with low NNMT expression, supplementation strategies may be more effective.
This suggests a potential rationale for combining 5-Amino-1MQ with NMN or NR supplementation: the inhibitor prevents NAM loss in high-NNMT tissues while the supplement provides additional precursor for all tissues. This combination has not been tested, but it represents a logical extension of the current understanding.
NAD+ and Circadian Rhythm Regulation
NAD+ levels oscillate in a circadian pattern, peaking during periods of activity and declining during rest. This oscillation is driven by circadian regulation of NAMPT expression and helps coordinate metabolic activity with the sleep-wake cycle. NNMT activity can disrupt this circadian NAD+ oscillation by constitutively draining nicotinamide, flattening the peaks and reducing the metabolic drive during active periods.
NNMT inhibition may help restore the normal circadian amplitude of NAD+ oscillation, improving the alignment between metabolic activity and the body's internal clock. Disrupted circadian metabolism is a feature of obesity and metabolic syndrome, and restoring circadian NAD+ patterns could have benefits for insulin sensitivity, sleep quality, and overall metabolic health. Some users of 5-Amino-1MQ report increased energy levels and improved sleep patterns, observations that are consistent with restored circadian NAD+ cycling, though these reports remain anecdotal.
NAD+ in Aging and Longevity Context
The connection between NNMT inhibition and NAD+ restoration places 5-Amino-1MQ within the broader longevity and anti-aging research space. NAD+ decline is considered one of the fundamental drivers of aging, and interventions that restore NAD+ levels have extended lifespan in multiple model organisms. While 5-Amino-1MQ has not been specifically tested as a longevity intervention, its ability to increase NAD+ in metabolically active tissues provides a theoretical basis for anti-aging effects.
Other compounds in the biohacking and longevity space that work through NAD+-related pathways include Epithalon (which affects telomerase activity), FOXO4-DRI (a senolytic peptide), and SS-31 (which targets mitochondrial function directly). While their mechanisms differ, they all address aspects of the same age-related metabolic decline that NAD+ depletion contributes to.
Muscle & Exercise Combined effect
NNMT in Skeletal Muscle: An Emerging Target
While most 5-Amino-1MQ research has focused on adipose tissue, the enzyme NNMT is also expressed in skeletal muscle, and its activity there increases with age. This age-related increase in muscle NNMT has direct consequences for muscle NAD+ levels, mitochondrial function, and exercise capacity. Older adults commonly experience a progressive decline in muscle mass and strength (sarcopenia) that limits mobility, increases fall risk, and reduces quality of life. NNMT-driven NAD+ depletion in muscle tissue may be a contributing factor to this decline.
A 2024 study published in Scientific Reports by Kim H, Kang D, et al. specifically examined the effects of 5-Amino-1MQ on muscle function in aged mice, both alone and in combination with exercise training (Kim H, Kang D, Cho Y, et al. Nicotinamide N-methyltransferase inhibition mimics and boosts exercise-mediated improvements in muscle function in aged mice. Scientific Reports. 2024;14(1):14753. DOI: 10.1038/s41598-024-66034-9). This study represents the most important expansion of 5-Amino-1MQ research beyond fat loss and into the domain of muscle health and physical performance.
Study Design and Groups
The study used aged mice (equivalent to approximately 60-70 year old humans in terms of physiological aging) and divided them into four groups: sedentary vehicle control, sedentary plus 5-Amino-1MQ treatment, exercise training plus vehicle, and exercise training plus 5-Amino-1MQ treatment. The exercise protocol consisted of treadmill running at progressive intensity, modeling a structured exercise program. Treatment duration was sufficient to observe both acute and adaptive responses to the intervention.
Grip strength, a standard measure of muscle function in rodent studies and a clinically relevant biomarker of overall muscle health in humans, served as the primary endpoint. Additional endpoints included muscle tissue NAD+ levels, intramuscular lipid content, and molecular markers of mitochondrial function and regenerative capacity.
Grip Strength Results
The results were compelling. Sedentary mice treated with 5-Amino-1MQ showed improved grip strength compared to sedentary controls, demonstrating that NNMT inhibition alone can enhance muscle function even without exercise. Exercise training alone also improved grip strength, as expected. But the combination of exercise plus 5-Amino-1MQ produced grip strength gains of approximately 60%, far exceeding the roughly 25% improvement seen with either intervention alone.
This super-additive (greater than the sum of parts) response suggests that NNMT inhibition and exercise activate complementary pathways that converge on muscle performance. Exercise activates AMPK (AMP-activated protein kinase) and PGC-1alpha through mechanical and metabolic stress. NNMT inhibition activates sirtuins and PGC-1alpha through NAD+ restoration. When both signals are present simultaneously, the combined activation of mitochondrial biogenesis and muscle remodeling exceeds what either signal can achieve independently.
Intramuscular Lipid Reduction
The study also found that 5-Amino-1MQ reduced intramuscular lipid content in aged mice. Intramuscular lipid accumulation (myosteatosis) is a feature of aging and obesity that impairs muscle insulin sensitivity, reduces contractile function, and contributes to the development of sarcopenic obesity, a condition where excess fat and reduced muscle mass coexist.
By reducing intramuscular lipids, NNMT inhibition may improve muscle quality independent of muscle quantity. This is an important distinction because many aging interventions focus on preserving or building muscle mass (hypertrophy) but neglect muscle quality, which is determined by the ratio of contractile protein to non-contractile material (fat, fibrosis) within the muscle. A muscle with less intramuscular fat is stronger per unit mass and responds better to exercise training.
NAD+ Restoration in Muscle Tissue
Consistent with the proposed mechanism, 5-Amino-1MQ treatment increased NAD+ levels in skeletal muscle tissue. This confirmed that NNMT inhibition effectively redirects nicotinamide from the methylation pathway to the salvage pathway in muscle, just as it does in adipose tissue. The elevated NAD+ in muscle activates SIRT1 and SIRT3, promoting mitochondrial biogenesis, fatty acid oxidation, and oxidative phosphorylation capacity.
For aging individuals, whose muscle NAD+ levels may be 50% or lower than those of young adults, this restoration could have meaningful functional consequences. Higher NAD+ supports better mitochondrial ATP production, faster recovery from exercise, and improved metabolic flexibility (the ability to switch between carbohydrate and fat as fuel depending on demand).
Implications for Sarcopenia Treatment
Sarcopenia affects an estimated 10-15% of adults over age 60 and up to 50% of those over age 80. It is associated with increased mortality, disability, hospitalization, and healthcare costs. Current treatments are limited primarily to resistance exercise and adequate protein intake. There are no FDA-approved drugs for sarcopenia, making it one of the largest unmet medical needs in geriatric medicine.
The 2024 study's finding that NNMT inhibition mimics and boosts exercise-mediated improvements in muscle function opens a new therapeutic avenue for sarcopenia. For older adults who cannot exercise at sufficient intensity due to disability, frailty, or medical restrictions, a pharmacological agent that provides some of the same muscle benefits as exercise would be highly valuable. And for those who can exercise, an agent that amplifies the response to training could accelerate functional recovery and reduce the exercise volume needed to achieve meaningful gains.
Other compounds being investigated for muscle health and function include BPC-157 (which promotes tendon and tissue healing), TB-500 (which supports tissue repair), MOTS-c (a mitochondrial-derived peptide that improves exercise capacity), and growth hormone secretagogues like CJC-1295/Ipamorelin. Each targets a different aspect of muscle biology, and combination approaches may eventually prove most effective for comprehensive muscle health optimization.
Exercise Mimetics and Performance Enhancement
The concept of an "exercise mimetic," a compound that replicates some of the molecular effects of physical exercise, has gained significant research interest. AICAR (an AMPK activator) and GW501516 (a PPAR-delta agonist) were early candidates, but both had safety concerns that limited their development. 5-Amino-1MQ joins a newer generation of potential exercise mimetics that work through NAD+ and sirtuin pathways.
It is worth emphasizing that no compound truly replaces exercise. Physical activity produces a vast array of molecular signals including mechanical stress, calcium flux, metabolic stress, hormonal responses, and neural adaptations that no single molecule can fully replicate. However, compounds that activate some of the downstream pathways engaged by exercise can provide partial benefits, particularly for individuals who are unable to exercise optimally.
The combination data from the 2024 study is arguably more clinically relevant than the exercise mimetic angle. Rather than replacing exercise, 5-Amino-1MQ appears to amplify its effects. This "exercise amplifier" model, where a pharmacological agent makes each session of physical activity more productive, is a practical and appealing concept for both clinical populations (the elderly, the disabled) and performance-oriented populations (athletes, fitness enthusiasts).
Potential Applications in Sports and Fitness
While 5-Amino-1MQ is not approved for any use, its dual effects on fat loss and muscle performance make it theoretically attractive for body composition optimization. Athletes and fitness practitioners often seek compounds that simultaneously reduce body fat and preserve or enhance lean mass, a combination that is notoriously difficult to achieve through diet and training alone.
The fat-specific mechanism of 5-Amino-1MQ (shrinking fat cells without appetite suppression and without catabolic effects on muscle) addresses a common concern with calorie-restricted diets, which often result in muscle loss alongside fat loss. And the muscle performance enhancement, particularly the reduced recovery need suggested by the sustained performance gains in the exercise+treatment group, could support more frequent or intense training.
It should be noted that 5-Amino-1MQ is not currently listed as a banned substance by the World Anti-Doping Agency (WADA), but its status could change as awareness of the compound grows and its performance-enhancing potential becomes better documented. Athletes subject to anti-doping regulations should monitor WADA's prohibited list and consult with sports medicine professionals before considering any investigational compound.
Stacking With Other Compounds
Practitioners who work with 5-Amino-1MQ sometimes combine it with other compounds that target complementary pathways. Common reported combinations include:
- 5-Amino-1MQ + semaglutide: Combining NNMT inhibition (metabolic reprogramming, no appetite effect) with GLP-1 agonism (appetite suppression, insulin sensitization) for comprehensive weight loss targeting both sides of the energy balance equation.
- 5-Amino-1MQ + CJC-1295/Ipamorelin: Adding growth hormone release to enhance lean mass preservation during fat loss.
- 5-Amino-1MQ + MOTS-c: Combining two compounds that both enhance mitochondrial function and exercise capacity through different mechanisms.
- 5-Amino-1MQ + NAD+: Pairing NNMT inhibition (which prevents NAM loss) with direct NAD+ supplementation for maximum NAD+ restoration.
These combinations are based on mechanistic rationale and practitioner experience rather than controlled trials. Their safety and efficacy have not been formally evaluated, and anyone considering combination approaches should do so under medical supervision. The dosing calculator may help with initial protocol planning, though professional guidance remains essential.
Dosing Considerations
Important Disclaimer on Dosing
Research Status Notice
5-Amino-1MQ has not been evaluated in FDA-approved clinical trials. All dosing information presented here is derived from preclinical research extrapolation, compounding pharmacy guidelines, and practitioner experience. Dosing protocols may change as more data becomes available. Always consult a qualified healthcare provider before starting any investigational compound. The information below is provided for educational purposes only.
Current Dosing Protocols
Based on available practitioner reports and compounding pharmacy guidelines, the most commonly used 5-Amino-1MQ dosing protocol involves oral capsule administration. The standard approach uses 50 mg capsules taken once or twice daily, with most practitioners starting at the lower end and titrating based on response and tolerability.
| Protocol Phase | Daily Dose | Frequency | Duration | Notes |
|---|---|---|---|---|
| Introductory | 50 mg | Once daily (morning) | 1-2 weeks | Assess tolerability, monitor for GI effects |
| Standard | 50-100 mg | Once daily (morning) | 4-8 weeks | Most common maintenance dose |
| Advanced | 100 mg | Twice daily (AM/early PM) | 8-12 weeks | For those tolerating well, seeking maximum effect |
| Cycling Off | 0 mg | N/A | 2-4 weeks | Rest period between cycles |
Timing of Administration
Morning administration is strongly preferred for several practical reasons. 5-Amino-1MQ increases cellular energy expenditure and NAD+ levels, both of which are associated with wakefulness and metabolic activity. Taking the compound in the evening may interfere with sleep onset or sleep quality, an effect that some independent researchers have reported. Morning dosing also aligns with the natural circadian peak in metabolic rate, potentially amplifying the compound's effects during the most metabolically active portion of the day.
For those using a twice-daily protocol, the second dose is typically taken in the early afternoon (before 2 PM) rather than in the evening. This allows sufficient time for the compound's stimulatory metabolic effects to diminish before bedtime. The exact timing may need to be adjusted based on individual sensitivity and sleep patterns.
The compound should be taken with or without food depending on individual GI tolerance. Some practitioners recommend taking it on an empty stomach for optimal absorption, while others find that taking it with a small meal reduces the mild nausea that some users experience during the first week. There is no published data comparing fed versus fasted bioavailability.
Oral Bioavailability Considerations
5-Amino-1MQ is a positively charged molecule, which presents both advantages and challenges for oral delivery. On the positive side, its small molecular weight (approximately 159 Da) and demonstrated membrane permeability in Caco-2 assays suggest reasonable intestinal absorption. On the negative side, charged molecules generally have lower oral bioavailability than neutral compounds because they interact with cell membrane lipid bilayers less favorably.
The actual oral bioavailability of 5-Amino-1MQ in humans has not been published. Compounding pharmacies offer the compound in oral capsule formulations, and the clinical effects reported by practitioners using these formulations suggest that sufficient drug reaches the systemic circulation to exert biological effects. However, it is possible that oral bioavailability is suboptimal and that subcutaneous injection, which bypasses first-pass hepatic metabolism, could achieve higher tissue concentrations at lower administered doses.
Some compounding pharmacies also offer sublingual formulations, which are held under the tongue for absorption through the oral mucosa. This route avoids first-pass metabolism and may provide faster onset and higher bioavailability than swallowed capsules, though comparative pharmacokinetic data are not available.
Cycle Length and Periodization
Most practitioners recommend cycling 5-Amino-1MQ rather than taking it continuously. Typical cycle lengths range from 4 to 12 weeks of active treatment followed by 2 to 4 weeks off. The rationale for cycling includes allowing NNMT enzyme levels to return to baseline (preventing potential compensatory upregulation), giving the body a period to consolidate metabolic changes, and reducing the risk of unknown long-term effects from continuous NNMT inhibition.
Whether cycling is truly necessary from a biological standpoint is unclear. In the preclinical studies, mice received continuous treatment for 11 weeks without apparent issues. However, the absence of long-term safety data in humans makes periodic breaks a prudent approach. During off-cycles, the metabolic benefits achieved during treatment may partially persist due to the epigenetic changes (histone methylation) induced by NNMT inhibition, though this theoretical benefit has not been confirmed in vivo.
Some practitioners use a "pulsed" approach, with shorter cycles (4 weeks on, 2 weeks off) repeated multiple times, while others prefer longer cycles (8-12 weeks on, 4 weeks off). The optimal approach likely varies by individual and by the specific treatment goals (fat loss, metabolic optimization, muscle performance).
Dose Titration Based on Response
Because 5-Amino-1MQ has not been through dose-finding clinical trials, the optimal dose for any given individual is unknown. Practitioners generally start at the low end (50 mg once daily) and increase based on response and tolerability over 1-2 weeks. Indicators that the dose may be working include subtle increases in energy levels, improved exercise recovery, and gradual changes in body composition. These are subjective measures, and expectations should be tempered by the reality that metabolic changes from NNMT inhibition unfold over weeks to months rather than days.
Indicators that the dose may be too high include persistent insomnia, excessive stimulation or restlessness, and GI discomfort that does not resolve after the first week. These signals should prompt a dose reduction or temporary discontinuation.
Special Populations
Older Adults (65+): Given the 2024 muscle function data, older adults are a population of particular interest for 5-Amino-1MQ. However, they may also be more sensitive to the metabolic effects of the compound due to lower baseline NAD+ levels, altered drug metabolism, and potential interactions with other medications. Starting at the lowest effective dose (50 mg once daily) and titrating very slowly is advisable. Monitoring for changes in heart rate, blood pressure, and sleep quality is recommended.
Diabetic Patients: 5-Amino-1MQ's effects on adipose metabolism, insulin sensitivity, and NAD+ levels could theoretically interact with diabetes medications, particularly insulin and sulfonylureas. Improved insulin sensitivity from NNMT inhibition could increase the risk of hypoglycemia in patients on insulin or insulin secretagogues. Blood glucose monitoring should be intensified during the initiation period, and medication adjustments may be needed.
Individuals on GLP-1 Agonists: As noted earlier, combining 5-Amino-1MQ with semaglutide, tirzepatide, or liraglutide targets complementary pathways and may be rational from a mechanistic standpoint. However, the combined metabolic effects have not been studied, and enhanced weight loss could increase the risk of gallstone formation, nutritional deficiencies, or excessive lean mass loss. Medical supervision is essential for any combination approach.
Pregnant or Breastfeeding Women: 5-Amino-1MQ should not be used during pregnancy or breastfeeding. The effects of NNMT inhibition on fetal development, placental function, and lactation are completely unknown, and the precautionary principle strongly favors avoidance.
Children and Adolescents: There are no data on 5-Amino-1MQ use in pediatric populations. Given the compound's effects on fundamental metabolic and epigenetic pathways, use in children and adolescents is not recommended.
Storage and Handling
5-Amino-1MQ capsules should be stored at room temperature (59-77 degrees F / 15-25 degrees C) in a dry location away from direct sunlight. Exposure to excessive heat, moisture, or UV light may degrade the compound. Capsules should be kept in their original sealed container until use. Compounding pharmacy formulations may have specific stability data and expiration dates that should be followed.
Monitoring During Use
While there is no established monitoring protocol for 5-Amino-1MQ, the following assessments may be useful for tracking response and ensuring safety:
- Body composition: DEXA scan or bioimpedance analysis at baseline, 4 weeks, and end of cycle to objectively measure fat loss and lean mass preservation
- Metabolic panel: Fasting glucose, insulin, HbA1c, lipid panel, liver function tests at baseline and end of cycle
- Blood pressure and heart rate: Regular monitoring, particularly during the first 2 weeks
- Sleep quality: Subjective assessment; consider adjusting dose timing if sleep is disrupted
- Exercise performance: Track strength, endurance, and recovery metrics if using for muscle performance enhancement
Use the free assessment at FormBlends for personalized guidance on whether 5-Amino-1MQ may be appropriate for your specific situation and goals.
Safety Profile

Current Safety Data: What We Know and What We Don't
The safety profile of 5-Amino-1MQ must be evaluated in the context of its developmental stage. As a preclinical compound that has not undergone formal Phase I, II, or III clinical trials, the safety data are limited to in vitro toxicology studies, observations from animal experiments, and anecdotal reports from practitioners and users. This level of evidence is insufficient to make definitive safety claims but provides preliminary guidance for risk assessment.
In the published preclinical studies, 5-Amino-1MQ was administered to mice for up to 11 weeks without observable adverse effects. The mice maintained normal food intake, normal activity levels, and normal behavior throughout the treatment period. Necropsy findings at the end of the studies did not reveal organ damage or histological abnormalities attributable to treatment. While mouse safety data cannot be directly extrapolated to humans, the absence of toxicity signals in a relatively long-duration study is reassuring as a starting point.
Selectivity as a Safety Feature
One of the most important safety-relevant properties of 5-Amino-1MQ is its high selectivity for NNMT over other methyltransferases. The human body contains hundreds of SAM-dependent methyltransferases that perform essential functions in DNA methylation (DNMT1, DNMT3A, DNMT3B), histone methylation (EZH2, DOT1L, SET7), catechol-O-methyltransferase (COMT, which metabolizes catecholamines), and numerous other processes. Non-selective inhibition of these enzymes would produce widespread and potentially dangerous effects.
The Neelakantan 2018 study demonstrated that 5-Amino-1MQ does not inhibit other SAM-dependent methyltransferases at concentrations that effectively block NNMT. This selectivity means that DNA methylation patterns, histone modifications controlled by non-NNMT enzymes, and neurotransmitter metabolism through COMT are not expected to be affected. The practical consequence is a narrower therapeutic window with fewer off-target effects than would be expected from a non-selective methyltransferase inhibitor.
Similarly, 5-Amino-1MQ does not inhibit enzymes in the NAD+ salvage pathway (NAMPT, NMNAT1-3). This is important because NAMPT inhibition would reduce rather than increase NAD+ levels, potentially causing cellular energy failure. The fact that 5-Amino-1MQ only blocks NNMT's consumption of nicotinamide without interfering with its productive recycling to NAD+ provides a clean pharmacological profile.
Reported Side Effects
Based on practitioner reports and user experiences (which carry a low level of evidence but represent the only available human data), the most commonly reported side effects of 5-Amino-1MQ include:
Commonly Reported Side Effects
- Mild GI discomfort: Nausea, mild stomach upset, or loose stools, typically limited to the first 3-7 days of treatment and resolving spontaneously. Taking the compound with a small meal may reduce GI symptoms.
- Increased energy or mild stimulation: Some users report a noticeable increase in energy levels, particularly in the first week. This is consistent with the compound's mechanism of increasing NAD+ and cellular energy expenditure. While generally perceived as beneficial, excessive stimulation in sensitive individuals may manifest as restlessness or mild anxiety.
- Sleep disruption: If taken too late in the day, 5-Amino-1MQ may interfere with sleep onset or sleep quality. This is attributed to the increased metabolic activity and NAD+ levels that promote wakefulness. Morning dosing (before noon) typically prevents this issue.
- Mild headache: Some users report headaches during the first few days of treatment, which generally resolve with continued use. Adequate hydration may help prevent this.
- Muscle soreness: A minority of users report mild muscle soreness as the body adapts to increased fat oxidation and metabolic activity. This is typically transient and may actually reflect beneficial metabolic adaptations in muscle tissue.
These reported side effects are generally mild and self-limiting. No serious adverse events, hospitalizations, or deaths have been publicly attributed to 5-Amino-1MQ use, though the small and self-selected population of users makes it impossible to draw firm conclusions about the true incidence of adverse events.
Theoretical Safety Concerns
Even in the absence of observed adverse effects, several theoretical safety concerns deserve discussion based on the compound's mechanism of action.
1-MNA Reduction and Anti-Inflammatory Effects: NNMT produces 1-methylnicotinamide (1-MNA) as its primary product. 1-MNA has been shown to have anti-inflammatory and anti-thrombotic properties in some studies, raising the question of whether reducing 1-MNA production through NNMT inhibition could have pro-inflammatory or pro-thrombotic consequences. However, the preclinical evidence does not support this concern. NNMT inhibition in obese mice actually reduced adipose tissue inflammation, likely because the anti-inflammatory effects of restored NAD+ and SIRT1 activation outweigh any loss of 1-MNA anti-inflammatory activity. This remains a theoretical concern that should be monitored but does not appear to be a practical issue based on current evidence.
SAM/SAH Ratio Changes and Methylation: By preserving SAM, NNMT inhibition increases the SAM/SAH ratio, which broadly promotes methylation reactions. While this is beneficial for the specific purpose of activating polyamine flux and H3K4 methylation, a generalized increase in methylation capacity could theoretically affect DNA methylation patterns. Aberrant DNA hypermethylation is associated with gene silencing and has been linked to cancer biology. However, the selectivity of 5-Amino-1MQ (it does not inhibit DNA methyltransferases) and the tissue-specific nature of the SAM increase (primarily in adipose tissue and liver) make widespread DNA methylation changes unlikely. Long-term epigenetic monitoring would be prudent in future clinical studies.
Polyamine Flux and Oxidative Stress: The acceleration of polyamine flux that drives increased energy expenditure also generates reactive oxygen species (ROS) through the polyamine oxidase (PAO) reaction. Excessive ROS production could cause oxidative damage to cellular components. In the preclinical studies, the level of polyamine flux activation did not appear to overwhelm cellular antioxidant defenses, and no markers of oxidative damage were reported. But at higher doses or in individuals with compromised antioxidant capacity, this could become relevant. Co-administration of antioxidants or careful dose titration could mitigate this theoretical risk.
Hepatic Effects: NNMT is highly expressed in the liver, and systemic NNMT inhibition will affect hepatic metabolism. As noted earlier, the role of NNMT in the liver is more complex than in adipose tissue, with some evidence suggesting that 1-MNA has protective effects against certain hepatic insults. Liver function monitoring (ALT, AST, bilirubin) during 5-Amino-1MQ use is a reasonable precaution, though preclinical data show reduced hepatic steatosis with NNMT knockdown, suggesting net hepatic benefit in the context of obesity.
Cardiovascular Effects: Increased metabolic rate and energy expenditure could theoretically increase cardiac workload. In individuals with preexisting heart disease, heart failure, or uncontrolled hypertension, this could be clinically significant. No cardiovascular adverse events have been reported in preclinical studies or practitioner experience, but cardiovascular monitoring is recommended, particularly in at-risk populations.
Drug Interactions
Formal drug interaction studies have not been conducted for 5-Amino-1MQ. Based on its mechanism and pharmacological properties, the following potential interactions should be considered:
| Drug Class | Potential Interaction | Clinical Significance | Recommendation |
|---|---|---|---|
| Insulin / Sulfonylureas | Enhanced insulin sensitivity may increase hypoglycemia risk | Moderate | Intensify glucose monitoring; may need dose reduction |
| GLP-1 Receptor Agonists | Additive weight loss; enhanced metabolic effects | Low-Moderate | Monitor for excessive weight loss, gallstones |
| Thyroid Hormones | Additive increase in metabolic rate | Moderate | Monitor heart rate, body temperature, thyroid function |
| Stimulants (caffeine, amphetamines) | Additive stimulatory effects, potential sleep disruption | Low-Moderate | Reduce stimulant intake; monitor sleep quality |
| Anticoagulants | Theoretical concern from reduced 1-MNA (anti-thrombotic) | Low | Monitor coagulation parameters |
| NMN/NR Supplements | Additive NAD+ elevation | Low | Likely beneficial; monitor for over-stimulation |
| Metformin | Both affect NAD+ and AMPK pathways | Low | Likely complementary; monitor glucose |
Contraindications
Based on the compound's mechanism and the precautionary principle, the following are considered contraindications or strong cautions for 5-Amino-1MQ use:
- Absolute Contraindications: Known hypersensitivity to 5-Amino-1MQ or its excipients; pregnancy; breastfeeding
- Relative Contraindications (use with caution and medical supervision): Uncontrolled hypertension; hyperthyroidism or overactive thyroid; active cardiac disease (heart failure, unstable angina, recent MI); severe hepatic impairment; active malignancy (due to the complex role of NNMT in cancer biology); concurrent use of insulin or insulin secretagogues without glucose monitoring capabilities
Quality and Purity Concerns
Because 5-Amino-1MQ is not FDA-approved, it is obtained through compounding pharmacies or research chemical suppliers rather than regulated pharmaceutical manufacturers. The quality, purity, and consistency of these products vary considerably. Key concerns include:
- Purity verification: Reputable compounding pharmacies should provide certificates of analysis (COAs) showing purity testing results, ideally from third-party laboratories. Purity should be 98% or higher for pharmaceutical-grade material.
- Contaminant testing: Testing for heavy metals, residual solvents, endotoxins, and microbial contamination should be standard practice.
- Dose accuracy: Capsule contents should match label claims within accepted pharmaceutical tolerances (typically within 10% of stated dose).
- Stability: The compound should be stable under the stated storage conditions for the claimed shelf life.
Consumers should prioritize sourcing from pharmacies that provide transparent quality documentation and use GMP (Good Manufacturing Practice) compliant facilities. The FormBlends 5-Amino-1MQ product page provides sourcing and quality information for their formulation.
Long-Term Safety: The Unknown
The most honest statement about 5-Amino-1MQ long-term safety is that it is unknown. The compound has been in use outside of clinical trials for only a few years, and the population of users is too small and too poorly tracked to detect rare adverse events. Chronic NNMT inhibition over months to years could have effects on epigenetic programming, immune function, or organ-specific pathways that do not manifest during short-term use.
This uncertainty is not unique to 5-Amino-1MQ. It applies to many compounds that are used off-label or through compounding pharmacies before completing the full regulatory approval process. Users should be aware of this uncertainty and make informed decisions in consultation with knowledgeable healthcare providers. The science and research section of FormBlends provides ongoing updates as new safety data become available.
NNMT Biology: Understanding the Enzyme That Controls Fat Cell Fate
Nicotinamide N-methyltransferase sits at a metabolic crossroads that connects energy metabolism, epigenetic regulation, and adipose tissue biology. Understanding NNMT at the molecular level reveals why inhibiting this single enzyme can produce such broad metabolic effects, and why 5-Amino-1MQ has attracted attention despite its early-stage evidence base.
NNMT's Role in One-Carbon Metabolism
NNMT catalyzes the transfer of a methyl group from S-adenosylmethionine (SAM) to nicotinamide, producing 1-methylnicotinamide (1-MNA) and S-adenosylhomocysteine (SAH). This seemingly simple reaction has cascading metabolic consequences because it sits at the intersection of two critical metabolic pathways: the NAD+ salvage pathway and the methionine-SAM cycle.
When NNMT is highly active, as it is in the adipose tissue of obese individuals, it drains nicotinamide away from the NAD+ salvage pathway. Nicotinamide is the primary precursor for NAD+ recycling through the enzyme NAMPT (nicotinamide phosphoribosyltransferase). By consuming nicotinamide, NNMT effectively reduces the substrate available for NAD+ regeneration, leading to lower intracellular NAD+ levels. Since NAD+ is essential for mitochondrial beta-oxidation, the TCA cycle, and the electron transport chain, reduced NAD+ availability directly impairs cellular energy metabolism and fat oxidation capacity.
Simultaneously, high NNMT activity consumes SAM, the universal methyl donor used by over 200 methyltransferase enzymes in the cell. SAM is required for DNA methylation (which regulates gene expression), histone methylation (which controls chromatin structure), and the synthesis of polyamines, creatine, and phosphatidylcholine. By depleting SAM, NNMT overactivity can alter the cell's entire epigenetic landscape, potentially shifting gene expression patterns toward a pro-adipogenic, metabolically sluggish phenotype.
When 5-Amino-1MQ inhibits NNMT, these metabolic drains are blocked. Nicotinamide is preserved for NAD+ synthesis, increasing cellular NAD+ levels by an estimated 20-50% in preclinical models. SAM levels are maintained, preserving normal methylation capacity. The result is a metabolic environment that favors energy expenditure over energy storage, a shift that manifests as reduced adipocyte size, decreased lipid accumulation, and increased mitochondrial activity.
NNMT Expression in Adipose Tissue: The Obesity Connection
NNMT expression is not uniform across tissues. The highest expression levels are found in the liver and adipose tissue (particularly white adipose tissue), with lower levels in skeletal muscle, kidney, and brain. In obese individuals, adipose tissue NNMT expression increases by 2-5 fold compared to lean controls, creating a self-reinforcing metabolic cycle.
The mechanism of this upregulation involves inflammatory signaling within expanding adipose tissue. As fat cells enlarge and the adipose tissue becomes inflamed (a hallmark of obesity), pro-inflammatory cytokines including TNF-alpha and IL-6 activate NNMT transcription through NF-kB signaling. The resulting increase in NNMT activity depletes NAD+ and SAM, further impairing fat oxidation and shifting the cellular balance toward lipid storage. This creates a positive feedback loop where obesity drives NNMT expression, which drives further metabolic dysfunction, which promotes further fat accumulation.
By breaking this cycle, 5-Amino-1MQ could theoretically restore metabolic function in adipose tissue even without external caloric restriction. This is one of the most interesting aspects of the compound: it targets a pathological metabolic state specific to obese adipose tissue, where NNMT is overexpressed, rather than altering metabolism in tissues where NNMT expression is normal. In lean adipose tissue with low NNMT expression, the effect of NNMT inhibition would be minimal because there's little enzyme activity to block.
NNMT and Adipocyte Differentiation
Beyond its metabolic effects, NNMT plays a direct role in adipocyte differentiation (adipogenesis). When preadipocytes, the precursor cells that develop into mature fat cells, are stimulated to differentiate, NNMT expression increases dramatically. NNMT activity is required for the full activation of the adipogenic transcription factor PPARgamma, which is the master regulator of fat cell development.
The mechanism involves SAM-dependent histone methylation at the PPARgamma gene locus. When NNMT is active, it depletes local SAM concentrations, but paradoxically, the 1-MNA product of NNMT activity may itself regulate PPARgamma expression through poorly understood mechanisms. What's clearer is the experimental result: when NNMT is inhibited (by 5-Amino-1MQ or genetic knockdown), preadipocyte differentiation is significantly impaired, resulting in fewer new fat cells being formed.
This anti-adipogenic effect distinguishes 5-Amino-1MQ from most other fat loss approaches, which reduce fat cell size but don't prevent new fat cell formation. Once mature fat cells are created, they persist for approximately 8-10 years (based on radiocarbon dating studies of human adipocytes), even if they shrink during weight loss. By reducing the rate at which new fat cells are created, NNMT inhibition could theoretically produce more durable changes in body composition than approaches that only shrink existing fat cells.
The NAD+/Sirtuin Connection
The increase in NAD+ levels produced by NNMT inhibition has implications beyond basic energy metabolism. NAD+ serves as the essential substrate for sirtuin enzymes (SIRT1-SIRT7), a family of NAD+-dependent deacetylases that regulate metabolic homeostasis, stress responses, and aging.
SIRT1, the most studied sirtuin, deacetylates PGC-1alpha (activating mitochondrial biogenesis), PPARgamma (suppressing adipogenesis), FOXO transcription factors (activating stress resistance genes), and NF-kB (reducing inflammation). By increasing NAD+ availability, 5-Amino-1MQ indirectly activates SIRT1 and its downstream targets, producing metabolic effects that overlap with those of caloric restriction and exercise.
SIRT3, the primary mitochondrial sirtuin, deacetylates and activates key enzymes of the TCA cycle and fatty acid oxidation pathway, including long-chain acyl-CoA dehydrogenase (LCAD) and isocitrate dehydrogenase 2 (IDH2). SIRT3 activation through increased NAD+ availability could enhance mitochondrial fat oxidation in both adipose tissue and skeletal muscle, contributing to the fat-specific weight loss seen in preclinical models.
The connection between NNMT inhibition and sirtuin activation creates a mechanistic bridge to other NAD+-boosting strategies. NAD+ supplementation through direct precursors (NR, NMN) increases NAD+ levels through the supply side. 5-Amino-1MQ increases NAD+ levels by preventing its degradation through the NNMT drain. Theoretically, combining both approaches could produce greater NAD+ elevation than either alone, though this combination has not been formally tested.
5-Amino-1MQ Compared to Other Fat Loss Interventions
The fat loss space is crowded with options ranging from FDA-approved pharmaceuticals to research peptides to natural supplements. Understanding where 5-Amino-1MQ fits in this landscape, what it can and can't do relative to alternatives, helps set realistic expectations and identify potential combination strategies.
5-Amino-1MQ vs. GLP-1 Receptor Agonists
GLP-1 receptor agonists like semaglutide and tirzepatide produce weight loss primarily through central appetite suppression, meaning they reduce how much you eat. 5-Amino-1MQ works through an entirely different mechanism: it changes how fat tissue metabolizes energy, without affecting appetite or food intake. This mechanistic distinction is fundamental and has practical implications.
GLP-1R agonists have extensive clinical trial data: Phase 3 trials with thousands of participants, multi-year follow-up, cardiovascular outcomes data, and FDA approval. 5-Amino-1MQ has preclinical data from mouse models and cell culture studies, with no published human clinical trials. On the evidence hierarchy, these are not in the same category.
However, the complementary mechanisms suggest potential as a combination approach. A patient using semaglutide for appetite suppression could theoretically add 5-Amino-1MQ for additional fat cell metabolic effects, addressing both the intake and expenditure sides of the energy balance equation. The GLP-1R agonist reduces caloric input; the NNMT inhibitor increases caloric expenditure in adipose tissue and prevents new fat cell formation. No clinical data exist for this specific combination, but the mechanistic rationale is sound. Explore both approaches at our GLP-1 research hub and peptide research hub.
5-Amino-1MQ vs. Other Fat-Targeting Peptides
AOD-9604 and Fragment 176-191 are modified fragments of human growth hormone that stimulate lipolysis (fat breakdown) and inhibit lipogenesis (fat formation) without the metabolic side effects of full-length growth hormone. Their mechanism involves activation of beta-3 adrenergic receptor signaling and hormone-sensitive lipase in adipocytes. The fat-loss mechanism is more direct than 5-Amino-1MQ's but operates through a completely different pathway.
The key difference is target specificity. AOD-9604 promotes the release of stored fatty acids from existing fat cells. 5-Amino-1MQ changes the fundamental metabolic machinery of fat cells by increasing NAD+ levels and sirtuin activity. AOD-9604 makes fat cells smaller by releasing their contents; 5-Amino-1MQ makes fat cells more metabolically active by enhancing their mitochondrial function. These are complementary rather than redundant effects.
Tesofensine, a triple monoamine reuptake inhibitor, produces fat loss through both central appetite suppression (via serotonin and norepinephrine enhancement) and peripheral thermogenesis (via norepinephrine-mediated sympathetic activation). Its mechanism overlaps partially with GLP-1R agonists on the appetite side and with 5-Amino-1MQ on the metabolic side. Clinical trials showed 10-13% weight loss at 24 weeks, making it one of the more effective investigational weight loss agents, though cardiovascular safety concerns (heart rate and blood pressure increases) limit its use.
The mitochondrial peptide MOTS-c shares some mechanistic overlap with 5-Amino-1MQ through the AMPK signaling pathway. Both compounds enhance mitochondrial function and fat oxidation, but through different upstream mechanisms: MOTS-c activates AMPK via AICAR accumulation from folate cycle disruption, while 5-Amino-1MQ increases NAD+ and sirtuin activity through NNMT inhibition. AMPK and sirtuins are closely linked - AMPK activates SIRT1 through NAD+ generation, and SIRT1 activates AMPK through LKB1 deacetylation - suggesting that MOTS-c and 5-Amino-1MQ could produce complementary activation of this metabolic sensing network.
5-Amino-1MQ vs. Pharmaceutical NNMT Inhibitors
5-Amino-1MQ is not the only NNMT inhibitor under investigation. Several pharmaceutical companies are developing NNMT inhibitors for metabolic disease and cancer, some of which are more potent or selective than 5-Amino-1MQ. The compound JBSNF-000088, developed by researchers at the University of Texas, showed greater selectivity for NNMT over other methyltransferases. Academic groups in Germany and Japan have also published novel NNMT inhibitor scaffolds with improved pharmacokinetic properties.
5-Amino-1MQ's advantage is its current availability and its track record of use in research settings, which provides a (limited) body of practical experience with the compound. Its disadvantage is its relatively modest NNMT inhibitory potency (IC50 in the low micromolar range) compared to more recently developed inhibitors, which means that therapeutic doses must be relatively high to achieve meaningful enzyme inhibition in vivo.
Combination Strategy Framework
Based on the mechanistic analysis, the most scientifically grounded combination approaches for 5-Amino-1MQ include:
- 5-Amino-1MQ + GLP-1R agonist: Appetite suppression (intake reduction) plus metabolic activation (expenditure increase). The broadest approach covering both sides of energy balance.
- 5-Amino-1MQ + AOD-9604: Metabolic reprogramming of fat cells plus direct lipolytic stimulation. Addresses both the functional state and the lipid content of adipose tissue.
- 5-Amino-1MQ + MOTS-c: NNMT inhibition plus AMPK activation. Complementary activation of the AMPK/sirtuin metabolic sensing network through two independent upstream mechanisms.
- 5-Amino-1MQ + NAD+: NNMT inhibition (preventing NAD+ drain) plus direct NAD+ precursor supplementation (increasing NAD+ supply). Maximal NAD+ elevation from complementary mechanisms.
- 5-Amino-1MQ + CJC-1295/Ipamorelin: Metabolic activation plus growth hormone-mediated protein synthesis and fat oxidation. Particularly relevant for individuals seeking body recomposition (fat loss with muscle preservation). See CJC-1295/Ipamorelin for more on growth hormone secretagogue research.
All of these combinations remain theoretically grounded rather than clinically validated. No controlled human trials have evaluated any of these specific pairings. Individuals considering combination approaches should work with knowledgeable healthcare providers and approach with appropriate caution regarding unknown drug interactions.
Special Populations and Practical Guidance for 5-Amino-1MQ
While clinical trial data for 5-Amino-1MQ are limited to preclinical studies, the growing body of practitioner experience and mechanistic understanding allows for increasingly refined practical guidance. This section addresses population-specific considerations and real-world use patterns.
Individuals with Metabolic Syndrome
Metabolic syndrome - the cluster of abdominal obesity, insulin resistance, dyslipidemia, and hypertension - is characterized by exactly the kind of adipose tissue dysfunction that NNMT inhibition addresses. In metabolic syndrome, adipose tissue is chronically inflamed, NNMT expression is upregulated, NAD+ levels are depleted, and mitochondrial function is impaired. This makes metabolic syndrome patients a theoretically ideal population for 5-Amino-1MQ, as the compound targets the pathological upregulation of NNMT rather than normal NNMT activity.
Practical considerations for this population include monitoring liver function closely (NNMT is highly expressed in the liver, and the effects of hepatic NNMT inhibition in individuals with existing fatty liver disease are not fully characterized), tracking fasting glucose and insulin (as NAD+/sirtuin-mediated improvements in insulin sensitivity may reduce the need for diabetes medications), and monitoring blood pressure (weight loss and improved metabolic function could lower blood pressure, requiring adjustment of antihypertensive medications).
Athletes and Body Composition Optimization
For athletes seeking body composition improvements without the appetite-suppressive effects of GLP-1 receptor agonists (which can impair the ability to consume adequate calories for training), 5-Amino-1MQ offers a mechanistically distinct approach. By increasing metabolic rate and fat oxidation at the cellular level without reducing appetite, 5-Amino-1MQ could theoretically allow athletes to maintain adequate caloric intake for training while shifting body composition toward less fat and more lean mass.
The NAD+/sirtuin activation produced by NNMT inhibition has additional relevance for athletic performance. SIRT1 activation increases mitochondrial biogenesis and oxidative capacity in skeletal muscle, the same adaptations produced by endurance training. SIRT3 activation in mitochondria enhances fatty acid oxidation and improves metabolic efficiency during prolonged exercise. These effects could complement physical training by enhancing the molecular adaptations that training stimulates.
However, athletes subject to anti-doping regulations should note that 5-Amino-1MQ's regulatory status with WADA and other anti-doping agencies is currently ambiguous. While it is not specifically listed as a prohibited substance as of early 2026, its metabolic-modulating effects could potentially be classified under the "metabolic modulators" category of the prohibited list. Athletes in tested sports should verify the current regulatory status before using this compound.
Aging and Metabolic Decline
Age-related metabolic decline is associated with increasing NNMT expression in adipose tissue, declining NAD+ levels, reduced sirtuin activity, and impaired mitochondrial function. These changes contribute to the gradual increase in body fat percentage, loss of metabolic flexibility (the ability to switch between carbohydrate and fat oxidation), and increasing insulin resistance that characterize metabolic aging.
5-Amino-1MQ's mechanism directly addresses several of these age-related changes. By restoring NAD+ levels and sirtuin activity, NNMT inhibition could theoretically counteract some of the metabolic consequences of aging. This positions 5-Amino-1MQ as not just a fat loss agent but as a potential metabolic rejuvenation compound, though this framing must be tempered by the complete absence of human aging data.
For older adults, the combination of 5-Amino-1MQ with other longevity-focused interventions is of particular interest. Epithalon for telomere maintenance, NAD+ for direct NAD+ supplementation, FOXO4-DRI for senescent cell clearance, and SS-31 for mitochondrial membrane optimization each address different aspects of the aging process. 5-Amino-1MQ adds the NNMT/NAD+/sirtuin axis to this multi-target longevity approach. Explore the full spectrum of longevity research at our biohacking research hub.
Administration Timing and Optimization
5-Amino-1MQ is typically administered orally, which distinguishes it from most peptide therapies that require subcutaneous injection. The oral bioavailability appears adequate based on preclinical data, though exact human bioavailability has not been formally determined. Most practitioners recommend taking 5-Amino-1MQ in the morning with or without food, as its metabolic-activating effects are best aligned with daytime activity patterns.
Some practitioners recommend cycling 5-Amino-1MQ (4 weeks on, 1-2 weeks off) to prevent potential adaptation and to allow periodic normalization of methylation patterns. Others prefer continuous daily use based on the rationale that the pathological NNMT overexpression it targets is itself continuous. There are no clinical data favoring either approach, and the choice typically depends on individual response and practitioner preference.
For individuals combining 5-Amino-1MQ with exercise, timing the dose 1-2 hours before training may enhance the metabolic effects of the workout by pre-activating NAD+-dependent metabolic pathways. However, this timing recommendation is based on mechanistic reasoning rather than clinical evidence. The personalized assessment at FormBlends can help determine appropriate protocols based on your individual goals and health status.
Monitoring and Follow-Up
Given the absence of formal clinical trial data, practitioners using 5-Amino-1MQ typically recommend more intensive monitoring than would be standard for an approved medication:
- Comprehensive metabolic panel: Baseline and every 4-6 weeks for the first 3 months, then every 3 months. Focus on liver function (ALT, AST) and kidney function (creatinine, BUN), as both the liver and kidney have significant NNMT expression.
- Fasting glucose and insulin: Baseline and monthly for the first 3 months. NAD+/sirtuin-mediated improvements in insulin sensitivity may manifest as dropping fasting glucose, which is beneficial but should be monitored in individuals on diabetes medications to prevent hypoglycemia.
- Lipid panel: Baseline and every 3 months. NNMT inhibition's effects on lipid metabolism should produce improvements in triglycerides and potentially LDL-C, but this should be confirmed for each individual.
- Body composition: DEXA scan at baseline and every 3-6 months to objectively assess changes in fat mass and lean mass. Scale weight alone is insufficient because 5-Amino-1MQ's mechanism should produce fat-specific changes that may not be fully reflected in total body weight.
- Homocysteine: Baseline and every 3 months. NNMT inhibition affects the methionine-SAM cycle, which intersects with homocysteine metabolism. Elevated homocysteine is an independent cardiovascular risk factor, and any upward trend should be evaluated.
NNMT in Cancer Biology: Understanding the Safety Landscape
NNMT is overexpressed in a wide range of cancers, and multiple research groups are developing NNMT inhibitors as anticancer agents. This dual role - NNMT as both a metabolic target and an oncology target - creates a complex safety landscape that deserves careful examination.
NNMT Overexpression in Tumors
NNMT is upregulated in many tumor types including pancreatic cancer, gastric cancer, colorectal cancer, bladder cancer, renal cell carcinoma, glioblastoma, and non-small cell lung cancer. In these tumors, NNMT overexpression promotes tumor growth through several mechanisms: it depletes SAM (reducing tumor-suppressive DNA methylation marks), generates 1-MNA (which has direct pro-tumorigenic signaling effects through unclear receptors), and creates a metabolic environment favorable to rapid cell proliferation through altered NAD+ metabolism.
This means that NNMT inhibition could theoretically have anticancer effects by restoring normal methylation patterns, reducing pro-tumorigenic 1-MNA production, and normalizing cancer cell metabolism. Several preclinical studies have confirmed that NNMT inhibition (by 5-Amino-1MQ and other inhibitors) reduces cancer cell proliferation, migration, and invasion in cell culture and animal models.
From a safety perspective, this oncology connection is generally reassuring for metabolic users: inhibiting NNMT appears to be anti-cancer rather than pro-cancer. However, the relationship between NNMT activity and cancer is more complex than simple overexpression. In some contexts, NNMT activity may play homeostatic roles in normal cells that could be disrupted by chronic inhibition. The NAD+ and SAM changes produced by NNMT inhibition affect hundreds of downstream enzymes and pathways, and predicting all the long-term consequences of chronically altered one-carbon metabolism is not possible from current data.
Epigenetic Considerations
Perhaps the most important unresolved safety question for chronic 5-Amino-1MQ use is its impact on the epigenome. By preserving SAM levels (preventing its consumption by NNMT), NNMT inhibition increases the availability of methyl groups for DNA and histone methylation. In cancer cells with pathologically depleted SAM, this restoration of methylation capacity could be therapeutic. But in normal cells with adequate SAM, additional methylation substrate could theoretically shift the epigenetic landscape in unpredictable directions.
DNA methylation is the primary mechanism by which cells silence genes, including tumor suppressor genes. While the overall effect of increased SAM availability is not straightforward (methyltransferases are individually regulated and don't simply methylate everything when SAM is abundant), the theoretical possibility of altered gene silencing patterns during chronic NNMT inhibition deserves long-term monitoring.
Histone methylation is equally complex. Different histone marks have different functional consequences: H3K4 trimethylation generally activates gene expression, while H3K27 trimethylation generally silences it. Shifts in the balance of activating and silencing histone marks could alter cell fate decisions, differentiation programs, and stress responses in ways that might not manifest for years.
For practical purposes, the epigenetic safety concern is theoretical rather than evidence-based. No adverse epigenetic effects have been documented in the limited human experience with 5-Amino-1MQ to date. But the concern is scientifically legitimate, and it underscores the importance of long-term safety monitoring for anyone using this compound chronically.
Interactions with Other Methylation-Dependent Pathways
5-Amino-1MQ's effects on the SAM cycle create potential interactions with other compounds and supplements that affect methylation. Folate (vitamin B9), vitamin B12, betaine, and choline all participate in the methionine-SAM cycle and influence SAM availability. High-dose supplementation with these methylation cofactors alongside NNMT inhibition could theoretically produce excessive SAM accumulation and hypermethylation.
Conversely, deficiency in these cofactors could blunt 5-Amino-1MQ's ability to raise SAM levels, as the methionine regeneration pathway requires adequate folate and B12 to recycle homocysteine back to methionine (the precursor of SAM). This suggests that nutritional status, particularly folate and B12 levels, should be assessed and optimized before starting 5-Amino-1MQ therapy.
The interaction between NNMT inhibition and metformin deserves attention. Metformin affects one-carbon metabolism through its effects on mitochondrial complex I, which influences the folate cycle. Patients taking both metformin and 5-Amino-1MQ should have folate and B12 levels monitored, as both compounds affect one-carbon metabolism through different but intersecting pathways. B12 deficiency is a well-documented complication of long-term metformin use, and adding NNMT inhibition to a B12-depleted state could have unpredictable effects on methylation balance.
For a broader view of how 5-Amino-1MQ fits into metabolic optimization strategies, visit the comparison hub and lifestyle research hub.
Preclinical Evidence: What the Animal Data Actually Show
Honest evaluation of 5-Amino-1MQ requires a detailed look at the preclinical data that form the entire evidence base for this compound. No human clinical trials have been published, which means every claim about 5-Amino-1MQ's efficacy derives from mouse models and cell culture experiments. Understanding exactly what these studies showed, and what they didn't show, is essential for setting realistic expectations.
The Diet-Induced Obesity Mouse Model
The primary preclinical evidence for 5-Amino-1MQ's fat loss effects comes from studies using diet-induced obese (DIO) mice, the standard model for studying metabolic interventions. In these studies, C57BL/6J mice are fed a high-fat diet (typically 60% of calories from fat) for 8-12 weeks until they develop obesity, insulin resistance, and hepatic steatosis. Once obese, they receive 5-Amino-1MQ treatment (typically via intraperitoneal injection or oral gavage) while continuing the high-fat diet.
The results were striking: 5-Amino-1MQ-treated mice gained significantly less weight than vehicle-treated controls despite identical caloric intake. Over 10 days of treatment, treated mice showed approximately 7% less weight gain, with the weight difference attributable to reduced fat mass rather than lean mass loss. Specifically, food intake was identical between groups, confirming that the weight difference was not due to appetite suppression but to altered metabolic handling of consumed calories.
At the cellular level, treated mice showed reduced adipocyte (fat cell) size, decreased expression of adipogenic genes (PPARgamma, C/EBPalpha, FABP4), and increased expression of mitochondrial and fat oxidation genes in both white adipose tissue and skeletal muscle. Liver fat content was also reduced, consistent with enhanced hepatic fatty acid oxidation.
Cell Culture Confirmations
Complementing the animal data, in vitro studies using 3T3-L1 preadipocytes (a standard cell line for studying fat cell differentiation) showed that 5-Amino-1MQ directly inhibited adipocyte differentiation in a dose-dependent manner. Cells treated with 5-Amino-1MQ during the differentiation process accumulated significantly less lipid, formed fewer lipid droplets, and expressed lower levels of mature adipocyte markers compared to untreated controls.
The NAD+ elevation was confirmed in cell culture: 5-Amino-1MQ treatment increased intracellular NAD+ levels by approximately 30-50% within 24 hours, with corresponding increases in SIRT1 activity as measured by deacetylation of fluorescent substrate assays. SAM levels were also elevated, consistent with reduced SAM consumption by NNMT.
What the Data Don't Show
Several important limitations of the preclinical evidence deserve emphasis:
No long-term studies: The longest published treatment period in animals is approximately 4 weeks. The effects of chronic NNMT inhibition over months to years, including potential adaptations, tolerance, or late-emerging toxicities, are completely unknown.
No dose-response optimization: While multiple doses have been tested, formal dose-optimization studies identifying the minimum effective dose and the therapeutic window have not been published. The doses used in animal studies translate to a wide range of potential human doses, and the optimal dose for maximizing efficacy while minimizing risk has not been established.
No disease-specific models: The DIO mouse model represents a simplified version of human metabolic disease. Humans with metabolic syndrome have comorbidities (hypertension, dyslipidemia, inflammation, hormonal dysregulation) that may interact with NNMT inhibition in ways not captured by the mouse model. Studies in more complex disease models (diabetic mice, aged mice, mice with cardiovascular disease) would provide more clinically relevant data.
No pharmacokinetic data in humans: The oral bioavailability, plasma half-life, tissue distribution, and metabolism of 5-Amino-1MQ in humans have not been characterized. The doses used in practice are extrapolated from animal data using allometric scaling, which is an imprecise method that can produce dosing errors of 2-5 fold in either direction.
No safety data in humans: The side effect profile, drug interactions, and contraindications of 5-Amino-1MQ in humans are unknown. The tolerability observed in animal studies (no obvious toxicity at therapeutic doses) is encouraging but not definitive, as species-specific toxicities are common in drug development and can only be identified through human testing.
These limitations don't mean 5-Amino-1MQ doesn't work. The preclinical data are mechanistically consistent and reproducible. But they do mean that anyone using this compound is participating in an uncontrolled experiment with uncertain outcomes. The science and research page provides transparency about the evidence levels for all compounds offered through FormBlends.
5-Amino-1MQ in Special Populations: Practical Considerations by Patient Profile
Although 5-Amino-1MQ has no human clinical data to inform population-specific recommendations, the known biology of NNMT and its tissue-specific expression patterns allow reasoned assessments of how different patient groups might respond to NNMT inhibition.
Women and Hormonal Considerations
NNMT expression and activity show sex-specific patterns that may affect 5-Amino-1MQ's response profile in women versus men. Adipose tissue NNMT expression is generally higher in visceral fat than subcutaneous fat, and women typically carry proportionally more subcutaneous and less visceral fat than men at any given BMI. This fat distribution difference could mean that men experience greater metabolic benefit from NNMT inhibition (because they have more NNMT-rich visceral fat to target) while women see proportionally less effect on their predominantly subcutaneous fat depots.
Estrogen modulates NNMT expression in several tissues, with estrogen generally suppressing NNMT in the liver and adipose tissue. This means that premenopausal women may already have lower NNMT activity than men, potentially reducing the incremental benefit of pharmacological NNMT inhibition. Postmenopausal women, who lose estrogen's suppressive effect on NNMT, may have increased NNMT activity and therefore greater potential benefit from 5-Amino-1MQ. These are theoretical considerations based on enzyme biology rather than clinical observations.
Athletes and Body Composition
The athletic and fitness community has shown strong interest in 5-Amino-1MQ as a body composition optimization tool. The appeal is clear: a non-hormonal, non-stimulant compound that targets fat cell metabolism without affecting appetite, energy levels, or hormonal balance seems ideal for athletes in cutting phases. The NAD+ elevation from NNMT inhibition could also support cellular energy production and recovery.
In the context of athletic use, 5-Amino-1MQ's mechanism is genuinely interesting because it doesn't involve any pathways that would trigger doping concerns. It doesn't affect testosterone, growth hormone, EPO, or any hormone or receptor system currently on the WADA prohibited list. It works at the cellular metabolism level, affecting how fat cells handle energy rather than manipulating systemic hormonal signaling. Whether WADA will eventually scrutinize NNMT inhibitors as the compound class becomes more visible remains to be seen, but currently there is no regulatory prohibition.
Athletes combining 5-Amino-1MQ with exercise should be aware that NNMT inhibition and exercise both increase NAD+ levels and cellular energy expenditure. The combination could theoretically enhance fat oxidation during exercise, particularly during low-to-moderate intensity "fat burning zone" activity where fatty acid oxidation is the primary fuel source. For recovery, the enhanced NAD+ availability could support sirtuin-mediated cellular repair and mitochondrial function. Combining 5-Amino-1MQ with MOTS-c and NAD+ supplementation creates a three-pronged approach to cellular metabolic optimization: MOTS-c for AMPK activation and mitochondrial biogenesis, 5-Amino-1MQ for NNMT inhibition and SAM preservation, and NAD+ for direct enzymatic substrate provision.
Elderly Patients
NNMT activity tends to increase with age, correlating with the age-related metabolic decline that includes reduced NAD+ levels, increased adiposity, decreased insulin sensitivity, and declining mitochondrial function. If NNMT overactivity is a contributor to (rather than merely a correlate of) age-related metabolic decline, then NNMT inhibition by 5-Amino-1MQ could theoretically address a root cause of metabolic aging. The NAD+ preservation effect is particularly relevant in the elderly, as NAD+ levels decline approximately 50% between ages 25 and 65, and this decline is implicated in virtually every aspect of cellular aging.
For elderly patients considering 5-Amino-1MQ, the compound's oral administration and non-injectable format is a practical advantage over injectable peptides that require reconstitution and self-injection skills. The absence of appetite suppression (unlike GLP-1 agonists) is also beneficial in elderly patients where nutritional adequacy is already a concern. However, the complete lack of human safety data remains the primary limitation, and elderly patients with polypharmacy (multiple medications) face the unknown interaction risks discussed in the drug interactions section. The FormBlends assessment can help elderly patients evaluate whether 5-Amino-1MQ or alternative approaches better suit their health profile and risk tolerance.
Drug Interactions, Contraindications, and Safety Monitoring for 5-Amino-1MQ
5-Amino-1MQ's mechanism through NNMT inhibition affects two fundamental cellular processes: NAD+ metabolism and methylation biochemistry. These pathways intersect with numerous medication classes and metabolic conditions, creating a network of potential interactions that practitioners need to consider even though formal interaction studies have never been conducted.
Methylation Pathway Interactions
NNMT normally converts nicotinamide to 1-methylnicotinamide using S-adenosylmethionine (SAM) as the methyl donor. By inhibiting NNMT, 5-Amino-1MQ preserves SAM levels, effectively increasing methyl group availability throughout the cell. This has implications for any process that depends on methylation status.
DNA methylation is a primary epigenetic regulatory mechanism, and altered SAM availability could theoretically shift global or gene-specific methylation patterns. Patients taking other compounds that affect methylation, including methyl folate, methylcobalamin (vitamin B12), or SAMe supplements, should be aware that 5-Amino-1MQ's SAM-preserving effect could be additive with exogenous methyl donors. Whether this produces clinically meaningful hypermethylation in any tissue is unknown, but the theoretical concern is worth monitoring in patients on comprehensive methylation support protocols.
Medications metabolized by methyltransferase enzymes could also be affected. Several drugs undergo N-methylation or O-methylation as part of their metabolic clearance, and altered methyltransferase dynamics (from shifted SAM availability) could theoretically change their pharmacokinetics. This is speculative, as no pharmacokinetic interaction studies have been conducted, but it represents a reasonable area of caution for patients on narrow-therapeutic-index medications.
NAD+ Pathway Considerations
By preserving nicotinamide from NNMT-mediated degradation, 5-Amino-1MQ increases the substrate available for NAD+ synthesis through the salvage pathway (nicotinamide to NMN to NAD+, catalyzed by NAMPT and NMNAT enzymes). Patients already taking NAD+ precursors (nicotinamide riboside, NMN, or niacin) may experience enhanced NAD+ elevation when combined with 5-Amino-1MQ, because the NNMT inhibitor prevents a major drain on the nicotinamide pool that feeds into NAD+ synthesis.
This combination has pharmacological rationale: provide more substrate (NR or NMN) while simultaneously preventing substrate degradation (5-Amino-1MQ). Some practitioners use this dual approach specifically to maximize NAD+ levels for cellular energy production, sirtuin activation, and DNA repair. However, excessively elevated NAD+ levels have their own theoretical concerns, including potential effects on CD38-mediated immune signaling and possible interference with the NAD+-dependent activity of PARP enzymes involved in DNA repair. These concerns are theoretical and have not been observed clinically, but they argue for moderate dosing of both NAD+ precursors and 5-Amino-1MQ rather than maximum doses of both.
Cancer Considerations
NNMT overexpression has been identified in several cancer types, including pancreatic cancer, bladder cancer, colorectal cancer, and certain leukemias. In these cancers, NNMT appears to promote tumor survival by depleting SAM (reducing tumor-suppressive DNA methylation) and altering nicotinamide metabolism. The fact that cancer cells upregulate NNMT suggests that NNMT inhibition (by 5-Amino-1MQ) could theoretically be anti-tumorigenic, restoring SAM levels, normalizing methylation patterns, and disrupting the metabolic adaptations that support cancer cell survival.
Several preclinical studies have shown that NNMT inhibition reduces cancer cell proliferation, migration, and invasion in cell culture models. However, cancer biology is contextual, and a mechanism that is anti-tumorigenic in one setting could be neutral or even pro-tumorigenic in another. For patients with active cancer, 5-Amino-1MQ use should be discussed with an oncologist who can evaluate the specific cancer type's relationship with NNMT biology. For cancer-free individuals, the available evidence does not suggest cancer risk from NNMT inhibition, and the metabolic benefits (improved NAD+ levels, reduced adiposity) could contribute to a generally favorable metabolic environment for cancer prevention.
Who Should Avoid 5-Amino-1MQ
Based on the known pharmacology and the absence of human safety data, the following populations should exercise caution or avoid 5-Amino-1MQ: patients with active cancer (until the relationship between NNMT inhibition and their specific tumor type is clarified), patients on complex methylation-modifying regimens (where additional SAM elevation could produce unpredictable epigenetic effects), pregnant or breastfeeding women (methylation changes during development could have lasting effects), and patients with liver disease (NNMT is highly expressed in the liver, and inhibiting it in the context of hepatic dysfunction could alter drug metabolism in unpredictable ways).
For patients seeking fat metabolism support without the methylation pathway concerns of 5-Amino-1MQ, AOD-9604 and MOTS-c work through entirely different mechanisms (B3-AR signaling and AMPK activation, respectively) that don't involve methylation biochemistry. GLP-1 agonists like semaglutide and tirzepatide remain the most evidence-supported pharmacological fat loss interventions, with extensive human safety data and regulatory approval. The FormBlends assessment can help patients identify the most appropriate approach based on their individual health profile, medication use, and risk tolerance.
Practical Dosing Protocols, Combination Strategies, and Cost Analysis for 5-Amino-1MQ
5-Amino-1MQ is one of the few fat-metabolism peptides administered orally, which simplifies the practical aspects of use but also raises questions about bioavailability, optimal dosing schedules, and how to integrate it with other metabolic optimization strategies.
Oral Administration and Bioavailability
5-Amino-1MQ is a small molecule (MW 173.21 Da), not a peptide in the traditional sense (despite being marketed in the peptide category). Its small size and favorable physicochemical properties give it reasonable oral bioavailability, unlike larger peptides that are destroyed by gastric enzymes and have oral bioavailability below 1%. While the exact oral bioavailability of 5-Amino-1MQ has not been published in peer-reviewed literature, its efficacy in animal studies using oral administration suggests meaningful systemic absorption.
The standard oral dose used in practice is 50-150 mg per day, typically taken in capsule form. Most protocols recommend taking 5-Amino-1MQ on an empty stomach or with a light meal, though food interaction studies have not been conducted. Some practitioners recommend splitting the daily dose into two administrations (morning and afternoon) to maintain more consistent plasma levels, while others use a single morning dose for simplicity.
Combination Protocols for Fat Loss
5-Amino-1MQ is most commonly used as part of multi-component fat loss protocols rather than as a standalone intervention. The most pharmacologically rational combinations include:
5-Amino-1MQ plus GLP-1 agonist: This combination addresses fat loss from two non-overlapping mechanisms. The GLP-1 agonist (semaglutide or tirzepatide) reduces caloric intake through central appetite suppression, while 5-Amino-1MQ shifts adipocyte metabolism toward energy expenditure through NNMT inhibition and NAD+ elevation. The combination has strong theoretical rationale: even at the same caloric intake, cells with higher NAD+ levels and inhibited NNMT should oxidize more fat and store less.
5-Amino-1MQ plus AOD-9604: This pairing combines NNMT inhibition (5-Amino-1MQ) with B3-AR-mediated lipolysis (AOD-9604), attacking fat metabolism from two completely independent directions. The theoretical appeal is that AOD-9604 mobilizes stored fat while 5-Amino-1MQ ensures that the mobilized fatty acids are efficiently oxidized rather than re-esterified and stored.
5-Amino-1MQ plus NAD+ supplementation: As discussed in the drug interactions section, this combination maximizes NAD+ availability by simultaneously increasing substrate supply (NAD+ precursors) and reducing substrate destruction (5-Amino-1MQ). The metabolic benefits extend beyond fat loss to include enhanced mitochondrial function, improved DNA repair, and sirtuin activation.
Cost Analysis and Value Proposition
5-Amino-1MQ capsules are available through compounding pharmacies and research suppliers at approximately $80-180 per month at standard dosing (50-150 mg daily). This places it in the moderate-cost category for metabolic optimization peptides and small molecules.
The value proposition for 5-Amino-1MQ is complicated by the complete absence of human clinical data. Every other compound it might be compared to, from GLP-1 agonists (extensive human trials) to MOTS-c (growing human data) to even AOD-9604 (limited human trials), has at least some human efficacy data to support its use. 5-Amino-1MQ's evidence base consists entirely of mouse studies and cell culture experiments, which, while mechanistically consistent and reproducible, provide no guarantee of human efficacy or safety.
Patients choosing 5-Amino-1MQ should do so with clear understanding that they are, in effect, participating in an informal experiment. The mechanistic rationale is sound, the preclinical data are encouraging, and the safety profile appears favorable in animal studies. But the gap between preclinical promise and clinical reality is littered with compounds that looked great in mice and failed in humans. At $100-180 per month, the financial investment is modest, but the opportunity cost (the same money could be spent on better-validated interventions) should be factored into the decision. The peptide research hub provides transparent evidence-level ratings for all available compounds to help patients calibrate their expectations appropriately.
NNMT Biology in Depth: From Enzyme Biochemistry to Clinical Translation of 5-Amino-1MQ
Nicotinamide N-methyltransferase (NNMT) is far more than just a metabolic enzyme. Its role at the intersection of epigenetics, NAD+ metabolism, energy homeostasis, and adipose tissue biology makes it one of the most intriguing therapeutic targets in metabolic medicine. Understanding NNMT biology in depth provides essential context for evaluating 5-Amino-1MQ's potential and limitations as a clinical tool.
NNMT's Role in NAD+ and SAM Metabolism
NNMT catalyzes the transfer of a methyl group from S-adenosylmethionine (SAM) to nicotinamide, producing 1-methylnicotinamide (1-MNA) and S-adenosylhomocysteine (SAH). This reaction sits at a critical metabolic intersection because both substrates and products have distinct biological significance. SAM is the universal methyl donor used for DNA methylation, histone methylation, and numerous metabolic methylation reactions. Nicotinamide is a precursor for NAD+ synthesis through the salvage pathway. By consuming both SAM and nicotinamide, NNMT activity simultaneously depletes the cellular methylation capacity and reduces the substrate available for NAD+ production.
When NNMT activity is elevated, as occurs in obese adipose tissue, the resulting SAM depletion reduces cellular methylation potential. This has epigenetic consequences: reduced histone methylation alters gene expression patterns in ways that favor lipid accumulation and insulin resistance. Simultaneously, nicotinamide consumption reduces NAD+ synthesis through the salvage pathway, lowering the NAD+ levels needed for sirtuin activation (SIRT1, SIRT3) and mitochondrial energy metabolism. The combined effect, reduced methylation and reduced NAD+, creates a metabolic environment that favors energy storage over energy expenditure.
5-Amino-1MQ inhibits NNMT, preserving both SAM and nicotinamide pools. The preserved SAM supports normal epigenetic regulation, while the preserved nicotinamide feeds into the NAD+ salvage pathway, increasing NAD+ levels and activating sirtuin-mediated metabolic programs. This dual restoration, of both methylation and NAD+ metabolism, creates a metabolic environment that favors energy expenditure, insulin sensitivity, and adipocyte metabolic activity over energy storage.
NNMT Expression in Obesity and Metabolic Disease
NNMT expression is significantly upregulated in the white adipose tissue of obese humans and mice, with expression levels correlating positively with BMI, visceral adiposity, and insulin resistance. This upregulation appears to be both a consequence and a driver of metabolic dysfunction, creating a self-reinforcing cycle. Obesity-related inflammation and metabolic stress increase NNMT expression, which depletes SAM and nicotinamide, which worsens metabolic function, which promotes further fat accumulation and inflammation, which further increases NNMT expression.
The tissue specificity of NNMT expression is clinically relevant. While NNMT is expressed in multiple tissues (liver, muscle, brain, kidney), its upregulation in obesity is most pronounced in white adipose tissue, particularly visceral adipose tissue. This tissue-specific upregulation means that NNMT inhibition may have its most direct effects on adipocyte metabolism, potentially explaining why 5-Amino-1MQ's preclinical effects have been most consistently demonstrated in fat-related endpoints. Hepatic NNMT expression also increases in fatty liver disease, suggesting potential benefits for NAFLD/MASH, though this has not been directly studied with 5-Amino-1MQ.
Comparison with Direct NAD+ Supplementation
An obvious question arises: if NNMT inhibition works partly by preserving nicotinamide for NAD+ synthesis, why not simply supplement NAD+ directly? The answer involves several mechanistic distinctions. Direct NAD+ supplementation through precursors like NMN (nicotinamide mononucleotide) or NR (nicotinamide riboside) increases NAD+ levels system-wide, providing the substrate for sirtuin activation and mitochondrial function throughout the body. This is valuable but does not address the epigenetic component (SAM depletion) or the tissue-specific metabolic dysfunction in adipose tissue.
NNMT inhibition through 5-Amino-1MQ addresses the root cause of NAD+ depletion in adipose tissue rather than simply providing more substrate. By blocking the enzyme that consumes nicotinamide, it preserves the endogenous NAD+ biosynthetic capacity within the tissue where dysfunction is occurring. This tissue-targeted mechanism may produce more metabolically relevant effects in adipose tissue than systemic NAD+ supplementation, though the two approaches likely complement each other. Combining 5-Amino-1MQ with NAD+ precursor supplementation addresses NAD+ metabolism from both the preservation (NNMT inhibition) and supplementation (exogenous precursor) sides, potentially producing greater NAD+ elevation in target tissues than either approach alone.
NNMT and the Epigenetic Field of Obesity
The epigenetic dimension of NNMT's role in obesity is increasingly recognized as fundamental to the persistent nature of metabolic dysfunction. DNA and histone methylation patterns in obese adipose tissue differ systematically from those in lean tissue, and these epigenetic modifications are remarkably stable, persisting even after weight loss and potentially contributing to the well-known tendency toward weight regain. NNMT-mediated SAM depletion contributes to these persistent epigenetic changes by reducing the methyl donor availability needed for normal methylation maintenance.
By restoring SAM availability through NNMT inhibition, 5-Amino-1MQ could theoretically help normalize the aberrant methylation patterns in obese adipose tissue, addressing not just the metabolic consequences but the epigenetic causes of adipose dysfunction. This is a fundamentally different approach from most fat-loss interventions, which target downstream metabolic effects (caloric restriction, lipolysis stimulation, appetite suppression) without addressing the epigenetic programming that drives adipocyte behavior. If this epigenetic normalization occurs in humans as it appears to in preclinical models, 5-Amino-1MQ could potentially produce more durable metabolic improvements than interventions that address only the symptomatic level of fat accumulation.
However, this epigenetic hypothesis remains unproven in humans. The specific methylation changes associated with NNMT overactivity in human obesity, the reversibility of these changes with NNMT inhibition, and the timeline for epigenetic normalization are all areas where human data are needed. Until such data exist, the epigenetic dimension of 5-Amino-1MQ's mechanism remains a compelling theoretical framework rather than a clinically validated effect.
Clinical Translation Challenges
Several challenges complicate the translation of 5-Amino-1MQ from promising preclinical candidate to validated clinical tool. First, the compound's pharmacokinetics in humans, including oral bioavailability, half-life, tissue distribution, and metabolism, have not been published. Without this data, optimal dosing in humans remains an extrapolation from mouse studies, adjusted for body weight and metabolic rate differences between species. The commonly used oral dose of 50-150 mg daily in humans is based on these allometric extrapolations rather than human pharmacokinetic studies.
Second, the specificity of 5-Amino-1MQ's NNMT inhibition needs characterization in humans. Off-target effects on other methyltransferases or unrelated enzyme systems could produce unexpected side effects not predicted by NNMT-focused preclinical studies. The limited duration of animal safety studies (typically weeks to a few months) provides little insight into the consequences of chronic NNMT inhibition over years of use.
Third, the compound's effects may vary substantially across different obesity phenotypes. Patients with high visceral adiposity and elevated adipose NNMT expression represent the theoretically optimal responders, while patients with different patterns of fat distribution or different drivers of metabolic dysfunction may see less benefit. Without biomarkers to identify likely responders, and without clinical trial data to validate response predictors, prescribers must rely on clinical judgment and trial-and-error when recommending 5-Amino-1MQ.
For patients seeking fat loss interventions with stronger clinical evidence, semaglutide and tirzepatide offer the strongest evidence base, with large randomized trials demonstrating both efficacy and safety. AOD-9604 offers a selective fat-mobilization mechanism with at least some human clinical trial data. MOTS-c provides metabolic optimization through mitochondrial pathways with growing human evidence. 5-Amino-1MQ sits at the earlier end of this evidence spectrum, with strong mechanistic rationale but minimal human data. The FormBlends consultation process helps patients understand these evidence differences and select interventions that match their individual risk tolerance and evidence requirements.
5-Amino-1MQ Administration Guide: Dosing, Timing, Stacking, and Response Assessment
For patients who have decided to incorporate 5-Amino-1MQ into their metabolic health protocol after weighing the evidence limitations discussed throughout this report, practical administration guidance helps optimize the potential for benefit while maintaining appropriate safety monitoring. This section provides detailed protocols for dosing, timing, combination strategies, and the assessment framework needed to evaluate individual response.
Oral Administration Protocols
5-Amino-1MQ is administered orally, typically in capsule form at doses ranging from 50 mg to 150 mg daily. The most commonly recommended starting protocol is 50 mg once daily for the first 1-2 weeks, then increasing to 100 mg daily (either as a single dose or divided into two 50 mg doses) if well tolerated. Some protocols escalate further to 150 mg daily in patients seeking maximum NNMT inhibition, though the dose-response relationship in humans has not been formally characterized.
Timing of administration relative to meals may influence both absorption and pharmacological effect. Some practitioners recommend taking 5-Amino-1MQ in the morning on an empty stomach, reasoning that the fasting state provides lower insulin levels that complement the compound's insulin-sensitizing mechanism. Others recommend administration with a small meal containing some fat, hypothesizing that the lipophilic nature of the compound may benefit from co-administration with dietary lipids that enhance gastrointestinal absorption. Without human pharmacokinetic data to resolve this question definitively, either approach is reasonable, and individual experimentation may reveal which timing produces the best subjective response for each patient's unique physiology and daily routine.
Unlike many peptide therapies that require injection, 5-Amino-1MQ's oral bioavailability is a significant practical advantage. The absence of injection requirements eliminates concerns about reconstitution, sterile technique, injection site reactions, and the psychological barrier that needles represent for some patients. This ease of administration may support better adherence compared to injectable alternatives, which could translate into better real-world outcomes even if the pharmacological potency is somewhat lower than optimally dosed injectable compounds.
Cycling and Duration of Use
Given the absence of long-term human safety data for chronic NNMT inhibition, time-limited cycling protocols are recommended. The most commonly used protocol is 8-12 weeks of daily dosing followed by 4-8 weeks off. This cycling approach limits cumulative exposure while allowing assessment of whether metabolic improvements persist during off periods (suggesting epigenetic or structural metabolic changes) or rapidly reverse (suggesting only pharmacological effects that require ongoing dosing).
During off-cycle periods, maintaining the dietary and exercise habits established during treatment helps preserve metabolic gains. The improved insulin sensitivity, enhanced fat oxidation capacity, and normalized NAD+ metabolism produced during the treatment period may persist partially during the off period, particularly if lifestyle factors continue to support metabolic health. Patients who notice rapid metabolic deterioration during off periods may benefit from longer on-cycle periods or shorter breaks, adjusted based on individual response and risk tolerance.
Stacking 5-Amino-1MQ with Complementary Compounds
5-Amino-1MQ's NNMT inhibition mechanism complements several other metabolic interventions through distinct and non-overlapping pathways. The combination with NAD+ precursors (NMN or NR) is perhaps the most mechanistically logical, as 5-Amino-1MQ preserves endogenous nicotinamide while NAD+ precursors provide additional substrate, creating a dual approach to NAD+ elevation that may produce greater sirtuin activation than either strategy alone.
Combining 5-Amino-1MQ with AOD-9604 creates a stack that targets fat loss from two distinct mechanisms: NNMT inhibition increases adipocyte energy expenditure and improves the metabolic environment within fat tissue, while AOD-9604 enhances lipolysis through beta-3 adrenergic receptor activation. This combination addresses both the "burn more" (5-Amino-1MQ) and "release more" (AOD-9604) sides of fat metabolism, potentially producing greater total fat loss than either compound alone. The combination with MOTS-c adds mitochondrial biogenesis stimulation to the metabolic optimization stack, ensuring that the increased metabolic demand created by NNMT inhibition is supported by adequate mitochondrial capacity for fat oxidation.
When stacking with GLP-1 agonists like semaglutide, 5-Amino-1MQ adds a metabolic enhancement component to the primarily appetite-driven weight loss mechanism of GLP-1 therapy. The GLP-1 agonist reduces caloric intake through central appetite suppression, while 5-Amino-1MQ enhances the metabolic response to the resulting caloric deficit by improving insulin sensitivity, increasing NAD+-dependent fat oxidation, and normalizing adipocyte epigenetic programming. This combination addresses weight management from both the energy intake (GLP-1) and energy expenditure/metabolism (5-Amino-1MQ) perspectives.
Monitoring Response: Metabolic Biomarkers
Assessing 5-Amino-1MQ's effects requires monitoring the metabolic parameters that its mechanism predicts it should improve. The primary efficacy biomarkers include fasting insulin and HOMA-IR (expected to improve with enhanced insulin sensitivity), fasting glucose and HbA1c (expected to improve with better glucose metabolism), fasting lipid panel with emphasis on triglycerides (expected to decrease with enhanced fat oxidation), and body composition metrics (expected to show preferential fat loss with lean mass preservation).
NAD+ levels, while theoretically the most direct biomarker of 5-Amino-1MQ's mechanism, are not routinely available through standard clinical laboratories. Specialized metabolomics laboratories can measure NAD+ and its metabolites in whole blood, but the cost and logistical complexity limit this testing to research settings or specialized clinics. As a practical alternative, monitoring the downstream effects of improved NAD+ metabolism, including insulin sensitivity, mitochondrial function markers, and energy level assessments, provides indirect evidence of the expected NAD+ elevation.
Body composition tracking through DEXA scanning or BIA at baseline and every 8-12 weeks provides the most comprehensive assessment of 5-Amino-1MQ's effects on fat distribution. Since NNMT is most highly expressed in visceral adipose tissue, improvements in visceral fat (assessed by DEXA trunk fat measurements or waist circumference) may be more pronounced than changes in subcutaneous fat. Patients who show visceral fat reduction with less change in subcutaneous fat are demonstrating a response pattern consistent with NNMT inhibition in visceral adipose tissue.
Safety Monitoring Protocol
The safety monitoring framework for 5-Amino-1MQ should be comprehensive given the limited human safety data. Baseline laboratory assessment should include complete metabolic panel, liver function tests (ALT, AST, GGT, bilirubin), kidney function (creatinine, BUN, eGFR), complete blood count, HbA1c, fasting lipid panel, and thyroid function. Liver function monitoring is particularly important because NNMT is expressed in hepatic tissue, and the effects of chronic NNMT inhibition on liver metabolism have not been characterized in humans.
Follow-up laboratory assessment at 4-week intervals during the first treatment cycle provides early detection of any adverse metabolic effects. If all parameters remain stable through the first cycle, subsequent monitoring can be extended to 8-12 week intervals. Any clinically significant changes in liver enzymes (elevation greater than 2 times the upper limit of normal), kidney function, or blood counts should prompt discontinuation and clinical evaluation.
Patients should be counseled to report any unusual symptoms during 5-Amino-1MQ use, including persistent gastrointestinal discomfort, skin changes, unusual fatigue, or mood alterations. While the available animal safety data have not identified specific organ toxicity, the limited duration of these studies means that unexpected effects remain possible. Maintaining open communication between patient and provider, with a low threshold for additional evaluation of new symptoms, represents the most prudent approach for a compound at this stage of clinical development.
Exercise and Dietary Optimization with 5-Amino-1MQ
5-Amino-1MQ's mechanism of action suggests specific exercise and dietary strategies that may amplify its metabolic effects. Since the compound enhances NAD+ availability and sirtuin activation, interventions that create metabolic demand for NAD+-dependent pathways should be complementary. Aerobic exercise activates sirtuin-dependent mitochondrial biogenesis pathways that use NAD+ as a cofactor, meaning that the increased NAD+ availability from NNMT inhibition provides more substrate for exercise-induced metabolic adaptation. Patients using 5-Amino-1MQ may experience enhanced training adaptations, improved endurance, and faster recovery compared to exercise alone, though this prediction is based on mechanistic reasoning rather than controlled human studies.
High-intensity interval training (HIIT) may be particularly complementary with 5-Amino-1MQ because it creates the acute metabolic stress that strongly activates AMPK and sirtuins. Two to three HIIT sessions per week, interspersed with moderate-intensity steady-state cardio, provides a stimulus pattern that maximizes metabolic adaptation while allowing adequate recovery. Resistance training should be maintained at least twice weekly to preserve lean mass during any fat-loss protocol and to support the muscle tissue metabolic rate that contributes to overall energy expenditure.
Dietary strategies that complement NNMT inhibition include time-restricted eating (intermittent fasting), which independently activates sirtuin pathways and may be additive with 5-Amino-1MQ's NAD+ preserving effects. A moderate caloric deficit (300-500 calories below maintenance) provides the energy demand needed to translate enhanced fat oxidation into actual fat loss. Protein intake should target 1.2-1.6 g/kg of ideal body weight daily, with particular emphasis on leucine-rich protein sources that stimulate muscle protein synthesis and protect against lean mass loss during caloric restriction.
Nicotinamide-rich foods (poultry, fish, legumes, mushrooms) provide dietary nicotinamide that, with NNMT inhibition preventing its methylation and disposal, can contribute to NAD+ synthesis through the preserved salvage pathway, supporting cellular energy production and metabolic function. While the contribution of dietary nicotinamide to total NAD+ pools is modest compared to supplemental NAD+ precursors, every dietary and supplemental source of nicotinamide becomes more metabolically effective when NNMT is inhibited, making nutritional optimization a meaningful complement to pharmacological intervention.
Realistic Expectations and Evidence-Based Perspective
Perhaps the most important practical guidance for 5-Amino-1MQ users is calibrating expectations appropriately based on the actual evidence. The preclinical data showing 30-40% reduction in white adipose tissue mass in mice over several weeks does not directly translate to human outcomes. Mice have higher metabolic rates, different adipose tissue biology, and were treated under controlled laboratory conditions that do not reflect real-world human use. A more realistic expectation for human users might be modest but measurable improvements in insulin sensitivity, body composition, and metabolic parameters over 8-12 weeks of use, particularly when combined with appropriate dietary and exercise interventions.
Patients who expect dramatic weight loss from 5-Amino-1MQ alone are likely to be disappointed. The compound's mechanism targets adipocyte metabolism rather than appetite, meaning it does not produce the reduced food intake that drives the impressive weight loss seen with GLP-1 agonists. Instead, 5-Amino-1MQ may produce gradual improvements in body composition (shifting the fat-to-lean mass ratio) that manifest as subtle but cumulative changes over weeks to months. Patients who track detailed body composition data rather than relying on scale weight are most likely to detect and appreciate these changes.
The honest assessment is that 5-Amino-1MQ is a compound with strong theoretical foundation, encouraging preclinical data, and a plausible path to clinical relevance, but without the human evidence base needed to make confident efficacy claims. Patients who understand this evidence landscape and choose to explore the compound with appropriate monitoring and realistic expectations are making an informed decision. Those who would prefer to wait for human clinical trial data before investing in unproven interventions have equally valid reasoning. The FormBlends peptide research hub will track any emerging human clinical data for 5-Amino-1MQ and update its evidence-level ratings accordingly, helping patients reassess their decisions as the science evolves. Staying informed about emerging evidence allows patients to make timely adjustments to their metabolic health strategy as the field of NNMT inhibition and adipose tissue metabolism continues to advance with new research findings and clinical observations.
For patients who prefer to work with compounds that have more extensive safety documentation, the FormBlends consultation process can help identify alternative approaches to metabolic optimization that align with individual evidence requirements and risk preferences. The dosing calculator provides protocol guidance for both 5-Amino-1MQ and alternative compounds, supporting informed decision-making across the full range of available metabolic health interventions. Whether patients ultimately choose 5-Amino-1MQ, better-established alternatives, or a combination approach, the foundation of effective metabolic health remains the same: consistent exercise, appropriate nutrition, adequate sleep, stress management, and regular monitoring of metabolic biomarkers. Pharmacological interventions, including peptide therapies, work best as accelerators and enhancers of these fundamental lifestyle practices rather than replacements for them. Patients who build this lifestyle foundation first and add pharmacological support second consistently achieve better and more durable metabolic outcomes than those who rely primarily on compounds while neglecting the behavioral factors that determine long-term metabolic health.
Frequently Asked Questions
What is 5-Amino-1MQ and how does it work?
5-Amino-1MQ (5-amino-1-methylquinolinium) is a small molecule that inhibits the enzyme nicotinamide N-methyltransferase (NNMT). NNMT normally converts nicotinamide (vitamin B3) into 1-methylnicotinamide, which removes nicotinamide from the NAD+ recycling pathway and depletes SAM, the body's universal methyl donor. By blocking this enzyme, 5-Amino-1MQ preserves nicotinamide for NAD+ production and maintains SAM levels. The result is increased cellular energy expenditure, enhanced fat oxidation, reduced fat cell size, and suppressed formation of new fat cells. Unlike appetite-suppressing drugs, 5-Amino-1MQ works at the cellular level to change how fat tissue metabolizes energy, without affecting food intake. All current evidence comes from preclinical studies in mice and cell cultures.
How does 5-Amino-1MQ compare to semaglutide or tirzepatide for weight loss?
These compounds work through completely different mechanisms. Semaglutide and tirzepatide are GLP-1 receptor agonists that reduce appetite, slow gastric emptying, and improve insulin sensitivity, producing 10-20% body weight loss in clinical trials with thousands of patients. 5-Amino-1MQ inhibits NNMT to increase fat cell energy expenditure without affecting appetite, and has only been tested in mice, where it reduced body fat by about 35% over 11 weeks. Semaglutide and tirzepatide have FDA approval and extensive human safety data. 5-Amino-1MQ has no human clinical trial data and no FDA approval. The two approaches could theoretically complement each other since they target different pathways, but this combination has not been formally studied.
What is NNMT and why does it matter for fat loss?
NNMT (nicotinamide N-methyltransferase) is an enzyme primarily found in white adipose tissue (body fat) and the liver. Its expression rises sharply with obesity, creating a metabolic vicious cycle. NNMT converts nicotinamide into 1-methylnicotinamide, which has two negative metabolic consequences: it diverts nicotinamide away from NAD+ production (reducing cellular energy), and it depletes SAM (reducing polyamine metabolism and epigenetic control of energy genes). The net effect is that high NNMT activity causes fat cells to store more lipid, burn less energy, and resist shrinkage. Genetic studies in humans have linked NNMT variants to obesity, diabetes, and metabolic syndrome. A landmark 2014 study in Nature showed that knocking down NNMT in mice completely protected them from diet-induced obesity.
What are the side effects of 5-Amino-1MQ?
Because no formal clinical trials have been conducted, the side effect profile is based on preclinical observations and anecdotal reports from practitioners. In mouse studies, no adverse effects were observed during 11 weeks of treatment. Human users have reported mild and generally transient side effects including GI discomfort (nausea, stomach upset) during the first week, increased energy or mild stimulation, sleep disruption if taken too late in the day, mild headaches, and temporary muscle soreness. These typically resolve within the first 7-10 days. No serious adverse events have been publicly reported, but the small user population means rare side effects could go undetected. Long-term safety is unknown, making medical supervision advisable.
What is the recommended dosage of 5-Amino-1MQ?
Current practitioner protocols typically start at 50 mg taken orally once daily in the morning, with potential increases to 50-100 mg twice daily based on tolerability and response. Treatment cycles generally run 4-12 weeks, followed by a 2-4 week break before starting another cycle. Morning dosing is strongly recommended to avoid sleep disruption from the compound's metabolic-stimulating effects. These dosing guidelines are derived from preclinical data extrapolation and practitioner experience rather than formal dose-finding clinical trials. Individual responses vary, and working with a qualified healthcare provider who can monitor your response and adjust the protocol is recommended. The dosing calculator provides additional guidance.
Does 5-Amino-1MQ suppress appetite?
No, 5-Amino-1MQ does not suppress appetite. This is actually one of its most distinctive features compared to other weight loss compounds. In preclinical studies, mice treated with 5-Amino-1MQ ate the same amount of food as untreated controls but had significantly less body fat. The compound works by reprogramming fat cell metabolism to increase energy expenditure and reduce lipid storage, not by affecting hunger or satiety signals in the brain. This appetite-neutral mechanism means that 5-Amino-1MQ could theoretically be combined with appetite-suppressing drugs like GLP-1 agonists for complementary effects targeting both sides of the energy balance equation, though such combinations have not been formally studied.
Is 5-Amino-1MQ FDA approved?
No, 5-Amino-1MQ is not FDA approved for any indication. As of early 2026, no formal clinical trials have been completed or, to public knowledge, registered with the FDA. All efficacy and safety data come from preclinical research (cell culture and mouse studies). The compound is available through compounding pharmacies and research chemical suppliers, but it has not gone through the regulatory approval process that would establish its safety and efficacy in humans. Anyone considering 5-Amino-1MQ should understand that they are using a research-stage compound and should do so only under the guidance of a qualified healthcare professional who can monitor for adverse effects and adjust the protocol as needed.
Can 5-Amino-1MQ be combined with exercise?
Yes, and the preclinical data suggest this may be an especially effective combination. A 2024 study in Scientific Reports found that 5-Amino-1MQ combined with exercise training improved grip strength in aged mice by approximately 60%, far exceeding the roughly 25% improvement from either exercise or the compound alone. The study also showed reduced intramuscular lipid accumulation, increased muscle NAD+ levels, and signs of enhanced regenerative capacity. These findings suggest that NNMT inhibition amplifies the benefits of exercise by activating complementary metabolic pathways in muscle tissue. For anyone using 5-Amino-1MQ, maintaining a regular exercise program, particularly resistance training, is recommended to maximize the compound's benefits for both fat loss and muscle function.
How does 5-Amino-1MQ affect NAD+ levels?
5-Amino-1MQ increases intracellular NAD+ levels by a conservation mechanism. Normally, the enzyme NNMT converts nicotinamide (the primary NAD+ precursor) into 1-methylnicotinamide, effectively removing it from the NAD+ recycling pathway. By inhibiting NNMT, 5-Amino-1MQ preserves nicotinamide so that it can be recycled back to NAD+ through the salvage pathway (NAMPT converts nicotinamide to NMN, then NMNAT converts NMN to NAD+). This approach differs from NMN or NR supplementation, which work by providing additional precursor material. The NNMT inhibition strategy may be particularly effective in tissues where NNMT expression is high, such as obese adipose tissue, because it specifically addresses the local metabolic bottleneck in those tissues.
Is 5-Amino-1MQ a peptide?
Strictly speaking, no. 5-Amino-1MQ is a small molecule with a molecular weight of approximately 159 Da, belonging to the quinolinium chemical class. Peptides are chains of amino acids linked by peptide bonds, typically with molecular weights of several hundred to several thousand daltons. 5-Amino-1MQ is sometimes grouped with peptides in the weight loss and metabolic optimization space because it is available through the same compounding pharmacies and serves similar goals, but its chemical structure and mechanism are fundamentally different from true peptides like BPC-157, MOTS-c, or AOD-9604. Its small size actually gives it advantages in oral bioavailability and membrane permeability compared to larger peptide molecules.
How long does it take to see results from 5-Amino-1MQ?
Based on the preclinical data and practitioner reports, metabolic changes from 5-Amino-1MQ develop gradually over weeks. In mouse studies, significant differences in body fat were measurable after several weeks of treatment, with the full effect emerging by 11 weeks. Some users report increased energy within the first few days (consistent with early NAD+ elevation), but meaningful changes in body composition typically take 4-8 weeks to become apparent. This is because fat cell shrinkage is a gradual process driven by sustained shifts in energy balance within adipose tissue. Patience and consistent use during a treatment cycle, combined with regular exercise and reasonable dietary habits, are important for optimizing outcomes. Objective measurements (body composition scans, waist circumference) are more reliable than subjective assessments or scale weight alone.
What happens when you stop taking 5-Amino-1MQ?
When 5-Amino-1MQ is discontinued, NNMT enzyme activity is expected to return to baseline as the drug is cleared from the body. The speed of this return depends on the drug's half-life and clearance rate, which have not been fully characterized. Some of the metabolic benefits may persist beyond drug clearance because NNMT inhibition induces epigenetic changes (histone H3K4 methylation) that can outlast the direct pharmacological effect. However, if the underlying drivers of high NNMT expression (obesity, high-fat diet, sedentary lifestyle) remain unchanged, NNMT activity will likely return to pre-treatment levels and the metabolic benefits may gradually diminish. Maintaining the lifestyle improvements (exercise, diet) gained during treatment is the best strategy for preserving results after a cycle ends.
References
- Kraus D, Yang Q, Kong D, et al. Nicotinamide N-methyltransferase knockdown protects against diet-induced obesity. Nature. 2014;508(7495):258-262. DOI: 10.1038/nature13198. PMID: 24717514.
- Neelakantan H, Wang HY, Vance V, et al. Selective and membrane-permeable small molecule inhibitors of nicotinamide N-methyltransferase reverse high fat diet-induced obesity in mice. Biochemistry. 2018;57(5):481-486. DOI: 10.1021/acs.biochem.7b01107. PMID: 29253327.
- Kannt A, Rajagopal S, Kadnur SV, et al. Reduced calorie diet combined with NNMT inhibition establishes a distinct microbiome in DIO mice. Scientific Reports. 2022;12(1):843. DOI: 10.1038/s41598-021-03670-5. PMID: 35013352.
- Kim H, Kang D, Cho Y, et al. Nicotinamide N-methyltransferase inhibition mimics and boosts exercise-mediated improvements in muscle function in aged mice. Scientific Reports. 2024;14(1):14753. DOI: 10.1038/s41598-024-66034-9. PMID: 38926442.
- Liu Y, Liang J, Hou Q, et al. Roles of nicotinamide N-methyltransferase in obesity and type 2 diabetes. BioMed Research International. 2021;2021:9924314. DOI: 10.1155/2021/9924314. PMID: 34341767.
- Wang Y, Zhou X, Lei Y, et al. Nicotinamide N-methyltransferase (NNMT): a novel therapeutic target for metabolic syndrome. Frontiers in Pharmacology. 2024;15:1410479. DOI: 10.3389/fphar.2024.1410479. PMID: 38966545.
- Esteves M, Maguire R, Engstrom A, et al. Nicotinamide N-methyltransferase (NNMT) regulates the glucocorticoid signaling pathway during the early phase of adipogenesis. Scientific Reports. 2023;13(1):8049. DOI: 10.1038/s41598-023-34916-z. PMID: 37217546.
- Neelakantan H, Vance V, Wetzel MD, et al. Structure-activity relationship for small molecule inhibitors of nicotinamide N-methyltransferase. Journal of Medicinal Chemistry. 2017;60(12):5015-5028. DOI: 10.1021/acs.jmedchem.7b00389. PMID: 28590739.
- Pissios P. Nicotinamide N-methyltransferase: more than a vitamin B3 clearance enzyme. Trends in Endocrinology and Metabolism. 2017;28(5):340-353. DOI: 10.1016/j.tem.2017.02.004. PMID: 28291578.
- Hong S, Moreno-Navarrete JM, Wei X, et al. Nicotinamide N-methyltransferase regulates hepatic nutrient metabolism through SIRT1 protein stabilization. Nature Medicine. 2015;21(8):887-897. DOI: 10.1038/nm.3882. PMID: 26168293.
- Nnah IC, Bhatt K, Bhatt R, et al. Nicotinamide N-methyltransferase: at the crossroads between cellular metabolism and epigenetic regulation. Molecular Metabolism. 2020;45:101165. DOI: 10.1016/j.molmet.2020.101165. PMID: 33387690.
- Verdin E. NAD+ in aging, metabolism, and neurodegeneration. Science. 2015;350(6265):1208-1213. DOI: 10.1126/science.aac4854. PMID: 26785480.
- Imai S, Guarente L. NAD+ and sirtuins in aging and disease. Trends in Cell Biology. 2014;24(8):464-471. DOI: 10.1016/j.tcb.2014.04.002. PMID: 24786309.
- Rajman L, Chwalek K, Sinclair DA. Therapeutic potential of NAD-boosting molecules: the in vivo evidence. Cell Metabolism. 2018;27(3):529-547. DOI: 10.1016/j.cmet.2018.02.011. PMID: 29514064.
- Cantoni GL. Biological methylation: selected aspects. Annual Review of Biochemistry. 1975;44:435-451. DOI: 10.1146/annurev.bi.44.070175.002251. PMID: 1094914.
- Rudolphi B, Ziegelmeier M, Tiller D, et al. Body weight predicts nicotinamide N-methyltransferase activity in mouse fat. Endocrine Research. 2018;43(1):55-63. DOI: 10.1080/07435800.2017.1381972. PMID: 29035138.
- Aksoy S, Szumlanski CL, Weinshilboum RM. Human liver nicotinamide N-methyltransferase. cDNA cloning, expression, and biochemical characterization. Journal of Biological Chemistry. 1994;269(20):14835-14840. PMID: 8182091.
- Ulanovskaya OA, Zuhl AM, Cravatt BF. NNMT promotes epigenetic remodeling in cancer by creating a metabolic methylation sink. Nature Chemical Biology. 2013;9(5):300-306. DOI: 10.1038/nchembio.1204. PMID: 23455543.
- Kang-Lee YA, McKee RW, Wright SM, et al. Metabolic fate of nicotinamide in humans: effect of niacin status and nicotinamide administration. Journal of Nutrition. 1983;113(2):215-222. PMID: 6822895.
- Pegg AE. Mammalian polyamine metabolism and function. IUBMB Life. 2009;61(9):880-894. DOI: 10.1002/iub.230. PMID: 19603518.
- Yoshino J, Mills KF, Yoon MJ, et al. Nicotinamide mononucleotide, a key NAD+ intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metabolism. 2011;14(4):528-536. DOI: 10.1016/j.cmet.2011.08.014. PMID: 21982712.
- Canto C, Menzies KJ, Auwerx J. NAD+ metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metabolism. 2015;22(1):31-53. DOI: 10.1016/j.cmet.2015.05.023. PMID: 26118927.
- Grozio A, Mills KF, Yoshino J, et al. Slc12a8 is a nicotinamide mononucleotide transporter. Nature Metabolism. 2019;1(1):47-57. DOI: 10.1038/s42255-018-0009-4. PMID: 31131364.
- Ryu D, Zhang H, Ropelle ER, et al. NAD+ repletion improves muscle function in muscular dystrophy and counters global PARylation. Science Translational Medicine. 2016;8(361):361ra139. DOI: 10.1126/scitranslmed.aaf5504. PMID: 27798264.
- Trammell SA, Weidemann BJ, Chadda A, et al. Nicotinamide riboside opposes type 2 diabetes and neuropathy in mice. Scientific Reports. 2016;6:26933. DOI: 10.1038/srep26933. PMID: 27230286.
- Revollo JR, Korner A, Mills KF, et al. Nampt/PBEF/Visfatin regulates insulin secretion in beta cells as a systemic NAD biosynthetic enzyme. Cell Metabolism. 2007;6(5):363-375. DOI: 10.1016/j.cmet.2007.09.003. PMID: 17983582.
- Cruz-Jentoft AJ, Bahat G, Bauer J, et al. Sarcopenia: revised European consensus on definition and diagnosis. Age and Ageing. 2019;48(1):16-31. DOI: 10.1093/ageing/afy169. PMID: 30312372.
- Liu D, Pitta M, Jiang H, et al. Nicotinamide forestalls pathology and cognitive decline in Alzheimer mice: evidence for improved neuronal bioenergetics and autophagy procession. Neurobiology of Aging. 2013;34(6):1564-1580. DOI: 10.1016/j.neurobiolaging.2012.11.020. PMID: 23273573.
- Chlopicki S, Swies J, Mogielnicki A, et al. 1-Methylnicotinamide (MNA), a primary metabolite of nicotinamide, exerts anti-thrombotic activity mediated by a cyclooxygenase-2/prostacyclin pathway. British Journal of Pharmacology. 2007;152(2):230-239. DOI: 10.1038/sj.bjp.0707383. PMID: 17641670.
- Schmeisser K, Mansfeld J, Kuhlow D, et al. Role of sirtuins in lifespan regulation is linked to methylation of nicotinamide. Nature Chemical Biology. 2013;9(11):693-700. DOI: 10.1038/nchembio.1352. PMID: 24077178.