Executive Summary
Key Takeaways
- Extended half-life: IGF-1 LR3's 20-30 hour half-life represents a roughly 100-fold increase over native IGF-1's 12-15 minute circulating duration, fundamentally changing its pharmacokinetic profile.
- Reduced IGFBP binding: The N-terminal extension and Glu3Arg substitution reduce IGFBP affinity by approximately 100-fold, leaving more free peptide available for receptor activation.
- Potency increase: In vitro and in vivo studies consistently demonstrate that LR3 is approximately 2-3 times more potent than native IGF-1 on a molar basis in stimulating cell proliferation and protein synthesis.
- Cancer concern: Large epidemiological studies, including UK Biobank analyses of over 300,000 participants, show positive associations between circulating IGF-1 levels and risk of breast cancer (HR=1.10), prostate cancer (HR=1.09), and colorectal cancer (HR=1.07).
- Hypoglycemia risk: As with recombinant IGF-1 (mecasermin), hypoglycemia remains the most common acute adverse effect, reported in approximately 42% of subjects receiving IGF-1 therapy in clinical trials.
IGF-1 LR3 (Long R3 Insulin-Like Growth Factor-1) is a synthetic, structurally modified analog of human IGF-1 that has been engineered to possess a dramatically extended biological half-life and greater potency than the endogenous hormone. With 83 amino acids compared to native IGF-1's 70, and a critical arginine-for-glutamic-acid substitution at position 3, this peptide variant evades sequestration by insulin-like growth factor binding proteins (IGFBPs), remaining bioavailable in circulation for 20 to 30 hours rather than the mere 12 to 15 minutes typical of unmodified IGF-1.
The insulin-like growth factor system sits at the center of some of the most consequential biology in human physiology. From fetal development to adult tissue maintenance, from skeletal muscle growth to wound healing, the IGF-1 receptor pathway influences cell proliferation, differentiation, protein synthesis, and programmed cell death. Understanding how IGF-1 LR3 interacts with this system requires appreciating both its therapeutic promise and its very real risks, particularly the relationship between sustained IGF-1 receptor activation and oncogenic potential.
This report provides a thorough examination of IGF-1 LR3 across seven critical dimensions: the fundamental biology of the IGF-1 signaling axis; the specific structural modifications that distinguish LR3 from the parent molecule; the preclinical and clinical evidence for muscle hypertrophy effects; how LR3 compares to other growth factor peptides including IGF-1 DES, growth hormone secretagogues like CJC-1295/Ipamorelin, and recombinant human IGF-1 (mecasermin); the safety profile with particular attention to cancer risk; and practical dosing considerations for researchers and clinicians.
Key Takeaways
- Extended half-life: IGF-1 LR3's 20-30 hour half-life represents a roughly 100-fold increase over native IGF-1's 12-15 minute circulating duration, fundamentally changing its pharmacokinetic profile.
- Reduced IGFBP binding: The N-terminal extension and Glu3Arg substitution reduce IGFBP affinity by approximately 100-fold, leaving more free peptide available for receptor activation.
- Potency increase: In vitro and in vivo studies consistently demonstrate that LR3 is approximately 2-3 times more potent than native IGF-1 on a molar basis in stimulating cell proliferation and protein synthesis.
- Cancer concern: Large epidemiological studies, including UK Biobank analyses of over 300,000 participants, show positive associations between circulating IGF-1 levels and risk of breast cancer (HR=1.10), prostate cancer (HR=1.09), and colorectal cancer (HR=1.07).
- Hypoglycemia risk: As with recombinant IGF-1 (mecasermin), hypoglycemia remains the most common acute adverse effect, reported in approximately 42% of subjects receiving IGF-1 therapy in clinical trials.
- Regulatory status: IGF-1 LR3 is not FDA-approved for any clinical indication. Mecasermin (Increlex), which uses unmodified recombinant IGF-1, is the only FDA-approved IGF-1 product, indicated specifically for severe primary IGF-1 deficiency in pediatric patients.
The research literature on IGF-1 LR3 spans decades of cell culture work, animal models, and observational data from the broader IGF-1 biology field. While no randomized controlled trials have been conducted specifically with LR3 in humans for muscle growth or anti-aging endpoints, the extensive mechanistic data and the clinical experience with mecasermin provide a substantial evidence base from which to draw conclusions about LR3's likely effects, limitations, and hazards.
What makes IGF-1 LR3 particularly interesting to researchers is that it essentially functions as a "constitutively active" version of IGF-1. By escaping the tight regulatory control that binding proteins normally exert over IGF-1 bioavailability, LR3 creates a pharmacological situation quite different from the pulsatile, tightly regulated IGF-1 signaling that occurs naturally. This distinction has profound implications for both efficacy and safety, and it forms the central tension running through every section of this report.
For those exploring the broader peptide research landscape, IGF-1 LR3 occupies a unique position at the intersection of growth hormone biology, anabolic signaling, and oncology. Its story illuminates fundamental questions about the tradeoffs between tissue growth promotion and the cellular proliferation that can, under certain circumstances, promote malignancy. Whether your interest is clinical, academic, or practical, a clear-eyed assessment of the evidence is essential before forming conclusions about this compound's appropriate role.
The sections that follow draw on peer-reviewed sources including publications in Nature Genetics, The Journal of Clinical Endocrinology & Metabolism, Cells, Skeletal Muscle, and Endocrine Reviews, among others. Every effort has been made to present the data accurately, cite primary sources, and distinguish between established findings and preliminary or extrapolated conclusions. The science and research section of FormBlends provides additional context on peptide pharmacology for readers seeking supplementary background.
How This Report Is Structured
Before proceeding to the detailed sections, a brief roadmap will help orient readers with different backgrounds and interests. Each section builds on the preceding material, but most can be read independently depending on your existing knowledge level.
The IGF-1 Biology section provides the foundational science, covering the GH-IGF-1 axis, receptor signaling cascades, the binding protein regulatory system, and IGF-1's roles across multiple organ systems. Readers with a strong background in endocrinology may want to skim this section, while those new to IGF-1 biology should read it thoroughly, as the concepts introduced here are essential for understanding the modification, safety, and dosing sections that follow.
The LR3 Modification and Extended Activity section details exactly what makes LR3 different from native IGF-1 at the molecular level, why these differences matter pharmacologically, and how the half-life comparison across IGF-1 variants shapes their respective risk-benefit profiles. This section includes the Chart.js visualization comparing half-lives across the three major IGF-1 variants.
The Muscle Hypertrophy Research section reviews the preclinical and clinical evidence for IGF-1's effects on skeletal muscle, including landmark transgenic animal studies, satellite cell biology, the myonuclear domain theory, fiber type specificity, and the critical interaction between exogenous IGF-1 and exercise-induced anabolic signaling. This is the longest section and the one most directly relevant to those considering LR3 for muscle growth purposes.
The Comparison section contextualizes LR3 relative to IGF-1 DES, growth hormone secretagogues, mecasermin, growth hormone, GLP-1 receptor agonists, and other peptides. This section is particularly useful for decision-making, as understanding the alternatives and their respective tradeoffs is essential for informed compound selection.
The Safety and Cancer Risk section is, in our assessment, the most important section of this report. It presents the epidemiological evidence linking IGF-1 to cancer risk, the lessons from acromegaly, the cardiovascular and metabolic concerns, and the emerging research that informs our understanding of IGF-1-related hazards. We encourage every reader to engage with this section thoroughly regardless of their intended use of the information.
The Dosing Considerations section provides practical information on dose ranges, administration technique, blood glucose management, reconstitution, storage, cycle structure, monitoring requirements, and post-cycle recovery. This section assumes readers have already absorbed the safety information and are seeking implementation guidance under medical supervision.
A Note on Evidence Quality
Throughout this report, we distinguish between different levels of evidence supporting various claims about IGF-1 LR3. The highest-quality evidence comes from randomized controlled trials in humans with mecasermin (which shares the same receptor target but different pharmacokinetics); these provide the most reliable data on IGF-1's effects and safety in clinical use. Transgenic animal models provide strong mechanistic evidence for specific biological effects but require cautious extrapolation to human pharmacology. In vitro (cell culture) studies provide detailed mechanistic insight but operate in simplified systems that may not reflect in vivo complexity. Epidemiological studies provide population-level risk associations but cannot establish causation for any individual. And anecdotal reports from the bodybuilding and research communities provide hypothesis-generating observations but are subject to placebo effects, recall bias, and confounding from concurrent use of other substances.
Where specific claims in this report are supported only by lower-quality evidence, we note this explicitly. Readers should calibrate their confidence accordingly and recognize that the absence of rigorous human clinical trial data for LR3 specifically means that much of what can be said about this compound rests on extrapolation from related compounds and mechanisms rather than direct observation.
Historical Context and Development Timeline
The discovery of insulin-like growth factors traces back to the 1950s, when researchers observed that sulfation of cartilage in response to growth hormone required an intermediary serum factor, initially called "sulfation factor" and later renamed "somatomedin." In 1978, Rinderknecht and Humbel purified and sequenced the two primary somatomedins from human serum, naming them IGF-1 and IGF-2 based on their structural similarity to proinsulin. This discovery established the fundamental paradigm of the GH-IGF axis that persists today.
The development of recombinant DNA technology in the 1980s made it possible to produce IGF-1 in quantities sufficient for research and, eventually, clinical use. Genentech and other biotechnology companies began exploring recombinant IGF-1 for growth disorders and metabolic conditions. The first human trials with recombinant IGF-1 began in the late 1980s, with Guler, Zapf, and Froesch publishing the first metabolic effects data in healthy adults in The New England Journal of Medicine in 1987, demonstrating the hormone's potent hypoglycemic activity.
The LR3 variant was developed in the late 1980s and early 1990s by researchers at GroPep Pty Ltd (later GroPep Bioreagents, now part of Novozymes) in Adelaide, Australia, led by Francis, Ross, Ballard, and colleagues. Their goal was to create an IGF-1 analog that could function effectively in cell culture media where binding proteins would otherwise sequester native IGF-1. The resulting Long R3 IGF-1 proved remarkably effective for this purpose and quickly became a standard cell culture supplement in the biopharmaceutical industry, where it remains commercially important today.
Mecasermin (brand name Increlex, manufactured by Ipsen) received FDA approval in 2005 for severe primary IGF-1 deficiency, marking the first and, to date, only regulatory approval of an IGF-1 product for therapeutic use. The approval was based on data from over 70 children treated in multicenter clinical trials demonstrating significant increases in height velocity and acceptable safety when administered with proper meal timing and monitoring.
The divergence between LR3's established role in biotechnology and its unapproved, experimental use in humans is an important contextual point. LR3 was never designed or tested for human therapeutic administration. Its adoption for muscle growth and anti-aging purposes emerged from the bodybuilding and performance enhancement communities in the early 2000s, driven by its commercial availability as a research reagent and its theoretical advantages over native IGF-1 (extended half-life, higher effective potency). This pattern of off-label migration from research tool to performance enhancer is common in the peptide space and creates significant safety uncertainties.
Understanding the Scope of This Report
This report is organized to serve readers with varying levels of background knowledge. The biology section provides a thorough foundation in IGF-1 physiology for those new to the topic. The LR3 modification section details the specific engineering that distinguishes this analog. The muscle hypertrophy section reviews the preclinical and clinical evidence base. The comparison section contextualizes LR3 relative to alternatives. The safety section provides an unflinching assessment of risks, with particular emphasis on the cancer question. And the dosing section offers practical considerations for those who, despite the risks, choose to proceed under medical supervision.
Throughout, we have endeavored to maintain a balance between accessibility and scientific rigor. Where evidence is strong, we say so clearly. Where it's preliminary, extrapolated, or based on animal models rather than human data, we note these limitations explicitly. The goal is to equip readers with the information needed to make informed decisions, not to advocate for or against IGF-1 LR3 use. The GLP-1 research hub and biohacking hub provide additional context on related compounds and optimization strategies for readers seeking a broader perspective.
IGF-1 Biology
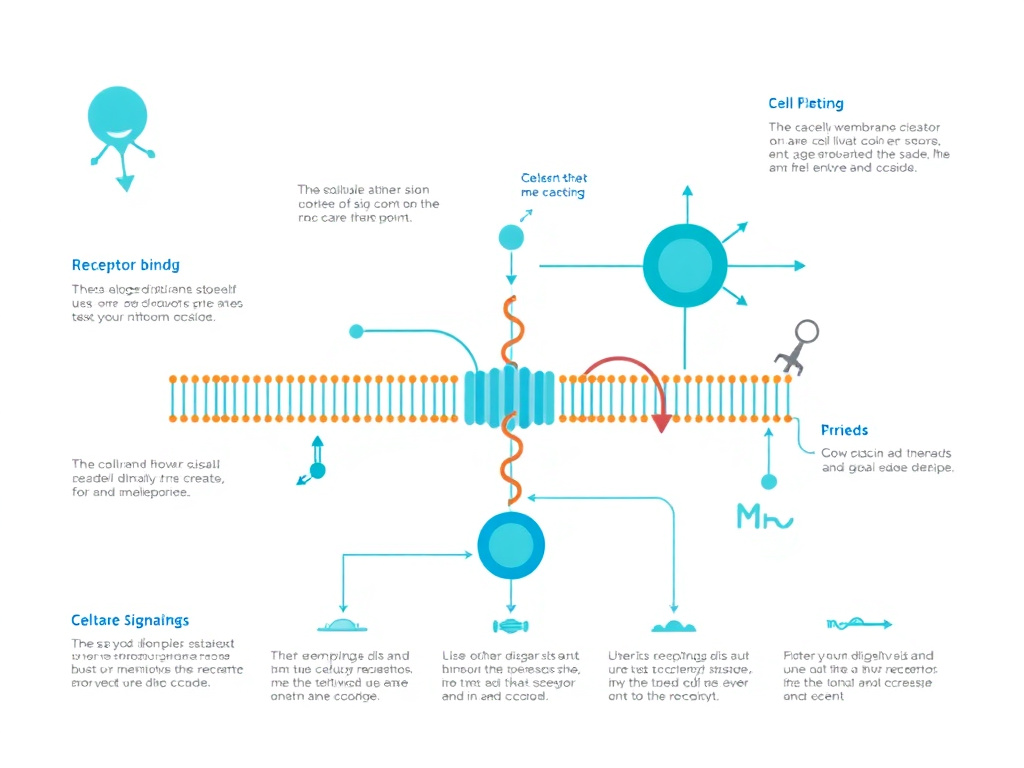
Figure 2: The IGF-1 receptor activates two major downstream signaling cascades: the PI3K/Akt/mTOR pathway (promoting protein synthesis) and the MAPK/ERK pathway (driving cell proliferation and differentiation).
What Is IGF-1 and Why Does It Matter?
Insulin-like growth factor 1 (IGF-1) is a 70-amino-acid polypeptide hormone that shares approximately 50% structural homology with insulin. Produced primarily in the liver under the stimulation of growth hormone (GH), IGF-1 also arises from local (autocrine and paracrine) production in virtually every tissue in the body, including skeletal muscle, bone, brain, kidney, and the gastrointestinal tract. It is one of the most potent natural activators of cell growth and survival signaling pathways, and its circulating concentration serves as a clinical biomarker for growth hormone status.
The name "insulin-like" reflects both structural similarity and functional overlap with insulin at supraphysiological concentrations. But the primary physiological role of IGF-1 is distinct from insulin's glucose-regulatory function. IGF-1's central purpose is to mediate the growth-promoting effects of growth hormone throughout postnatal life, driving longitudinal bone growth during childhood and adolescence, maintaining lean tissue mass in adulthood, and supporting tissue repair processes throughout the lifespan.
Circulating IGF-1 levels fluctuate across the lifespan in a characteristic pattern. Levels are relatively low during early childhood, rise sharply during puberty (peaking around age 14-16), and then decline progressively throughout adult life. By age 60, IGF-1 concentrations are typically 50-60% lower than peak pubertal values. This age-related decline has attracted significant attention from researchers studying sarcopenia (age-related muscle loss) and has fueled interest in IGF-1-based interventions, including compounds like IGF-1 LR3, for their potential to counteract muscle wasting.
The GH-IGF-1 Axis
The growth hormone-IGF-1 axis represents one of the body's primary anabolic signaling systems. Growth hormone, secreted in pulsatile bursts from the anterior pituitary gland (predominantly during deep sleep and in response to exercise), travels to the liver and other tissues where it activates the JAK2/STAT5 signaling pathway. This triggers transcription of the IGF-1 gene, leading to synthesis and secretion of IGF-1 into the bloodstream.
This axis operates under classical negative feedback control. Circulating IGF-1 acts back on the hypothalamus and pituitary to suppress GH release, creating a self-regulating loop. Growth hormone releasing hormone (GHRH) from the hypothalamus stimulates GH secretion, while somatostatin inhibits it. Peptides like sermorelin and CJC-1295/Ipamorelin work by stimulating this axis at the hypothalamic-pituitary level, increasing endogenous GH and consequently IGF-1 production.
The clinical significance of this axis is well illustrated by two extremes. GH excess, as seen in acromegaly from pituitary adenomas, produces chronically elevated IGF-1 that drives visceral organ enlargement, soft tissue hypertrophy, insulin resistance, cardiomyopathy, and increased cancer risk. Without treatment, acromegaly reduces life expectancy by roughly 30%. At the other extreme, severe GH or IGF-1 deficiency produces dwarfism in children and contributes to reduced lean mass, increased adiposity, impaired bone density, and decreased quality of life in adults.
The IGF-1 Receptor and Downstream Signaling
IGF-1 exerts its biological effects primarily through the type 1 IGF receptor (IGF-1R), a transmembrane tyrosine kinase receptor structurally related to the insulin receptor. The IGF-1R exists as a preformed dimer on the cell surface. When IGF-1 binds to the extracellular alpha subunits, it triggers autophosphorylation of tyrosine residues on the intracellular beta subunits, creating docking sites for adapter proteins - most critically, insulin receptor substrate proteins (IRS-1 and IRS-2).
From this initial receptor activation, two major signaling cascades diverge:
The PI3K/Akt/mTOR Pathway. Phosphorylated IRS recruits and activates phosphatidylinositol-3-kinase (PI3K), which generates the lipid second messenger PIP3 from PIP2 at the cell membrane. PIP3 recruits both PDK1 and Akt (also called protein kinase B) to the membrane, where PDK1 phosphorylates and activates Akt. Activated Akt then orchestrates a broad program of anabolic signaling:
- Protein synthesis activation: Akt phosphorylates and activates mTORC1 (mammalian target of rapamycin complex 1), which in turn activates p70S6 kinase and inhibits 4E-BP1, stimulating ribosomal protein translation and cap-dependent mRNA translation. Akt also inactivates GSK3-beta, relieving inhibition of the translation initiation factor eIF2B.
- Protein degradation suppression: Akt phosphorylates and inactivates FoxO transcription factors (FoxO1, FoxO3a, FoxO4), preventing their nuclear translocation. This suppresses transcription of the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1, which are the primary drivers of ubiquitin-proteasome-mediated muscle protein breakdown.
- Anti-apoptotic signaling: Akt phosphorylates and inactivates the pro-apoptotic protein Bad, while also activating NF-kB through IKK phosphorylation. These signals promote cell survival.
The MAPK/ERK Pathway. Simultaneously, IGF-1R activation recruits the adapter protein Shc, which binds Grb2 and the guanine nucleotide exchange factor SOS. This activates the small GTPase Ras, initiating the Raf/MEK/ERK kinase cascade. The MAPK/ERK pathway primarily drives cell proliferation and differentiation rather than the protein synthesis/anti-atrophy program controlled by PI3K/Akt. In skeletal muscle, ERK activation promotes satellite cell proliferation, a critical step in muscle repair and hypertrophy that requires the donation of new myonuclei from activated satellite cells to growing or damaged myofibers.
The Binding Protein System: Nature's Regulatory Layer
Perhaps the most important aspect of IGF-1 biology for understanding LR3 is the insulin-like growth factor binding protein (IGFBP) system. Six high-affinity IGFBPs (IGFBP-1 through IGFBP-6) regulate virtually every aspect of IGF-1 bioavailability, distribution, and activity. In healthy adults, approximately 98-99% of circulating IGF-1 is bound to IGFBPs at any given moment, leaving only 1-2% as "free" IGF-1 capable of engaging the receptor.
The total circulating IGF-1 concentration in healthy adults is approximately 100 nM (nanomolar). Yet the IGF-1 receptor on most cell types approaches saturation at concentrations of just 5 nM or lower. This enormous excess of total IGF-1 relative to receptor capacity makes clear that IGFBPs don't merely serve as passive carriers - they are the primary regulators of IGF-1 biological activity. Without binding proteins, the IGF-1 system would be constitutively activated at maximum levels, a situation that the disease acromegaly approximates and that carries severe health consequences.
IGFBP-3 is the dominant carrier, accounting for roughly 75-80% of circulating IGF-1 binding. It forms a ternary complex with IGF-1 and a third protein called acid-labile subunit (ALS). This 150 kDa ternary complex is too large to cross capillary barriers, effectively trapping IGF-1 in the vascular compartment and extending its circulating half-life from approximately 10-12 minutes (free) to 12-15 hours (in the ternary complex). To reach target tissues, the complex must partially dissociate, a process facilitated by specific IGFBP proteases that cleave the binding proteins and release bioactive IGF-1 locally.
IGFBP-5 deserves special mention because of its dual role. It can both inhibit IGF-1 action (by sequestering ligand away from the receptor) and potentiate it (by concentrating IGF-1 at specific tissue sites and delivering it to the receptor surface). IGFBP-5 is the most evolutionarily conserved of the six binding proteins, suggesting its regulatory functions are under strong selective pressure. In skeletal muscle, IGFBP-5 expression increases during differentiation and is thought to play an important role in fine-tuning IGF-1's effects on myogenesis.
Local vs. Systemic IGF-1: Different Roles, Different Risks
A critical distinction in IGF-1 biology is between endocrine (liver-derived, circulating) IGF-1 and autocrine/paracrine (locally produced) IGF-1. Liver-specific IGF-1 gene knockout mice maintain normal postnatal growth despite a 75% reduction in circulating IGF-1, demonstrating that local tissue production can largely substitute for systemic delivery in supporting growth.
In skeletal muscle, locally produced IGF-1 appears to play the dominant role in exercise-induced hypertrophy. Mechanical loading triggers expression of specific IGF-1 splice variants, including mechano-growth factor (MGF, also known as IGF-1Ec), which activates satellite cells and initiates the repair/growth response. This local production operates independently of the GH-liver axis, which is why muscle growth can occur even in states of relative GH deficiency (such as during caloric restriction combined with resistance exercise).
This distinction matters for understanding IGF-1 LR3 because exogenous administration creates a systemic exposure pattern fundamentally different from the tightly regulated, pulsatile, locally produced IGF-1 that tissues normally encounter. The consequences of this difference are explored in detail in the safety section of this report. Readers interested in the broader context of growth hormone peptides may find our peptide research hub helpful for understanding how different compounds in this class interact with the GH-IGF-1 axis.
IGF-1 Beyond Muscle: The Multi-System Hormone
While muscle growth applications receive the most attention in the peptide research community, IGF-1 influences virtually every organ system:
- Bone: IGF-1 stimulates both osteoblast (bone-forming cell) proliferation and differentiation, and plays a critical role in longitudinal bone growth through effects on the growth plate. Low IGF-1 is associated with osteoporosis risk.
- Brain: IGF-1 crosses the blood-brain barrier and promotes neuronal survival, synaptogenesis, and myelination. It supports hippocampal neurogenesis and has been investigated in Alzheimer's disease models. A 2024 study demonstrated that intranasal LR3-IGF-1 treatment promoted amyloid plaque remodeling in cerebral cortex of 5XFAD mice, with enhanced microglial uptake of amyloid-beta peptide in vitro (PMID: 39610283).
- Heart: IGF-1 promotes physiological cardiac hypertrophy (the beneficial kind associated with exercise) and protects cardiomyocytes from apoptosis. However, chronic excess (as in acromegaly) causes pathological remodeling.
- Immune system: IGF-1 supports T-cell development in the thymus and modulates immune cell function. Related peptides like Thymosin Alpha-1 also influence immune regulation through complementary pathways.
- Wound healing: IGF-1 accelerates wound closure by promoting fibroblast proliferation, collagen synthesis, and keratinocyte migration. Peptides like BPC-157 and TB-500 share some of these tissue repair properties through different mechanisms.
- Metabolism: IGF-1 has insulin-like effects on glucose uptake and can cause hypoglycemia at supraphysiological levels. It also influences lipid metabolism and adipocyte differentiation.
This broad biological footprint explains both the appeal and the risk profile of IGF-1-based compounds. The same receptor activation that promotes muscle growth also influences cancer cell proliferation, organ size, and metabolic homeostasis. Any intervention that modifies IGF-1 signaling has the potential to produce far-reaching effects beyond the intended target tissue.
IGF-1 Gene Structure and Splice Variants
The human IGF-1 gene is located on chromosome 12q23.2 and spans approximately 85 kilobases. Its transcription is remarkably complex, with multiple promoters, alternative splicing of the 5' and 3' untranslated regions, and differential exon usage generating at least six distinct mRNA transcripts. All mature IGF-1 protein is identical regardless of transcript variant (the 70-amino-acid mature peptide), but the propeptide and signal peptide sequences differ, affecting tissue-specific expression patterns and post-translational processing.
Three IGF-1 splice variants have received particular attention in the muscle biology literature:
- IGF-1Ea (systemic/liver isoform): The predominant circulating form produced by the liver under GH stimulation. This is the main endocrine form of IGF-1 and the basis for mecasermin.
- IGF-1Eb: A splice variant expressed in various tissues with distinct post-translational processing characteristics.
- IGF-1Ec (Mechano-Growth Factor, MGF): A splice variant specifically upregulated in skeletal muscle following mechanical loading (exercise). MGF contains a unique C-terminal E peptide that appears to independently activate satellite cells. The initial pulse of MGF expression following muscle damage is thought to be a key signal for satellite cell activation, distinct from the subsequent sustained expression of IGF-1Ea that supports proliferation and differentiation.
The significance of these splice variants for understanding LR3 is indirect but important. In normal physiology, muscle growth signaling involves a carefully orchestrated temporal sequence: an initial MGF pulse activates satellite cells, followed by sustained IGF-1Ea expression that drives their proliferation, differentiation, and fusion. LR3 administration bypasses this temporal programming entirely, providing a constant, high-level IGF-1R activation that may produce different downstream effects on satellite cell dynamics than the natural sequential signaling pattern.
The Insulin/IGF-1 Receptor Family
Understanding the full receptor family is important for appreciating LR3's pharmacology. Three closely related receptors exist:
- Type 1 IGF Receptor (IGF-1R): The primary receptor for IGF-1. A heterotetrameric transmembrane tyrosine kinase (two alpha and two beta subunits). Mediates most of IGF-1's growth-promoting, anti-apoptotic, and anabolic effects.
- Insulin Receptor (IR): Structurally homologous to IGF-1R (approximately 60% amino acid identity in the kinase domain). Exists in two isoforms: IR-A (expressed in fetal tissues and many cancers; binds IGF-2 with high affinity) and IR-B (the classic adult metabolic insulin receptor). IGF-1 can bind IR at supraphysiological concentrations, contributing to its insulin-like metabolic effects.
- Hybrid Receptors (IGF-1R/IR): Because IGF-1R and IR are both dimeric receptors assembled from half-receptors, cells expressing both can form hybrid receptors containing one IGF-1R half and one IR half. These hybrids preferentially bind IGF-1 over insulin and signal predominantly through IGF-1R-type pathways. They are abundant in skeletal muscle and may be an important mediator of IGF-1's muscle-specific effects.
The type 2 IGF receptor (IGF-2R, also known as the mannose-6-phosphate receptor) is structurally unrelated and does not activate intracellular signaling. It primarily functions as a clearance receptor, binding and internalizing IGF-2 for degradation. It has very low affinity for IGF-1 and is not a significant target for LR3.
IGF-1 in the Context of the Somatotropic Axis
The broader somatotropic (growth) axis involves multiple interacting components beyond the simple GH-to-IGF-1 pathway. Ghrelin, produced primarily by the stomach, stimulates GH release through the growth hormone secretagogue receptor (GHSR). This is the mechanism exploited by MK-677 (Ibutamoren) and GHRP-6. GHRH from the hypothalamus acts on pituitary somatotrophs through the GHRH receptor, which is the target of sermorelin, tesamorelin, and CJC-1295 DAC. Somatostatin, also from the hypothalamus, inhibits GH release and provides the "off switch" for GH pulsatility.
The interplay between these regulatory inputs creates the characteristic pulsatile GH secretion pattern: 6-12 discrete GH pulses per 24 hours in young adults, with the largest pulse occurring during the first period of deep (slow-wave) sleep. This pulsatility is not merely a consequence of the regulatory architecture; it is functionally important. Continuous GH infusion at the same total daily dose as pulsatile administration produces inferior IGF-1 induction and different metabolic effects, demonstrating that the temporal pattern of stimulation matters for the biological outcome.
This context helps explain why directly administering IGF-1 (or LR3) is pharmacologically very different from stimulating the endogenous axis with secretagogues. The secretagogue approach preserves the architecture of pulsatile regulation, negative feedback, and tissue-specific IGF-1 production. Direct IGF-1 administration overrides all of these regulatory mechanisms, creating a sustained, uniform receptor activation state that no part of the natural system was designed to handle.
Species Considerations in IGF-1 Research
Much of the IGF-1 research relevant to LR3 was conducted in rodent models, and species differences in IGF-1 biology deserve acknowledgment. Human and rodent IGF-1 share high sequence homology (the mature proteins differ at only a few positions), and the receptor systems are well-conserved. However, there are meaningful differences in IGFBP expression patterns, circulating IGF-1 concentrations, and the relative contributions of hepatic versus local IGF-1 production.
Mice and rats have approximately 10-fold higher circulating IGF-1 concentrations relative to body mass than humans, and their IGFBP profiles differ, particularly in the relative abundance of IGFBP-1 and IGFBP-4. These differences mean that the effective "free fraction" of IGF-1 - and consequently the degree of receptor activation - may not translate linearly between species. Dose extrapolation from rodent studies to human use requires careful allometric scaling and should account for differences in IGFBP milieu.
The guinea pig studies by Lok and colleagues using LR3 directly are somewhat more translatable because guinea pigs, unlike mice and rats, share certain features of human IGF-1 metabolism, including sensitivity to GH-axis suppression by exogenous IGF-1. However, even guinea pig data requires cautious interpretation when applied to human dosing decisions.
The Laron Syndrome Natural Experiment
One of the most informative natural experiments in IGF-1 biology comes from studies of Laron syndrome, a genetic condition characterized by growth hormone receptor insensitivity that results in extremely low circulating IGF-1 despite elevated GH levels. Laron syndrome patients are very short (adult height typically 120-130 cm) but have an extraordinary resistance to cancer.
Guevara-Aguirre and colleagues (2011) studied a cohort of 99 Laron syndrome subjects in Ecuador over more than two decades and found that not a single case of cancer occurred in the IGF-1-deficient group, compared to a 17% cancer incidence in unaffected relatives. The group also showed dramatically reduced diabetes prevalence despite obesity. These findings, published in Science Translational Medicine, provided some of the strongest human evidence linking low IGF-1 signaling to cancer protection and metabolic resilience (Guevara-Aguirre J, Balasubramanian P, Guevara-Aguirre M, et al. Growth hormone receptor deficiency is associated with a major reduction in pro-aging signaling, cancer, and diabetes in humans. Sci Transl Med. 2011;3(70):70ra13. PMID: 21325617).
More recently, long-term follow-up of this cohort and similar GH receptor-deficient populations in Israel and other countries has confirmed and extended these findings. The Ecuadorian cohort members who are heterozygous carriers (one functional copy of the GH receptor gene) have normal stature but intermediate IGF-1 levels and intermediate cancer risk, suggesting a dose-response relationship between IGF-1 signaling capacity and cancer susceptibility across the full range of human genetic variation.
The Laron syndrome data provides a powerful counterpoint to the muscle growth narrative around IGF-1. While IGF-1 is clearly necessary for normal growth and tissue maintenance, the complete absence of cancer in a population with lifelong IGF-1 deficiency suggests that the IGF-1 signaling pathway is not merely associated with cancer risk but may be causally required for cancer development in many contexts. This has obvious implications for anyone contemplating exogenous elevation of IGF-1 signaling through compounds like LR3.
LR3 Modification & Extended Activity
What Is IGF-1 LR3 and How Does It Differ from Native IGF-1?
IGF-1 LR3, formally known as Long R3 Insulin-Like Growth Factor-1, is a recombinant protein analog of human IGF-1 that incorporates two specific structural modifications designed to evade the binding protein regulatory system while retaining full agonist activity at the IGF-1 receptor. The result is a molecule with dramatically altered pharmacokinetics but largely preserved pharmacodynamics relative to the parent compound.
The first modification is an N-terminal extension consisting of 13 additional amino acids (a methionine followed by a sequence derived from synthetic design). This extension increases the total length of the protein from 70 amino acids (native IGF-1) to 83 amino acids, adding approximately 1.4 kDa to the molecular weight. The N-terminal region of IGF-1 is critically involved in IGFBP binding, and the addition of this extension peptide sterically hinders the interaction between the modified protein and the binding protein family.
The second modification is a single point substitution: the glutamic acid (Glu) normally present at position 3 of the mature IGF-1 sequence is replaced with arginine (Arg). This Glu3Arg substitution, from which the "R3" in the name derives, further disrupts the electrostatic and structural complementarity required for high-affinity IGFBP binding. The glutamic acid at position 3 sits within a key contact region for IGFBP-3 and IGFBP-5, and its replacement with the positively charged arginine introduces a charge reversal that weakens the interaction substantially.
Quantifying the Binding Protein Escape
The combined effect of these two modifications reduces IGFBP affinity by roughly 100-fold compared to native IGF-1. To appreciate why this matters, consider the numbers. In normal physiology, approximately 99% of circulating IGF-1 is bound to IGFBPs and biologically inactive at any given moment. Only the 1% "free fraction" can engage the IGF-1 receptor. When you administer IGF-1 LR3, the binding protein system can only sequester a small fraction of the dose. The vast majority remains as free, receptor-available peptide.
This has two immediate consequences:
Extended half-life. Native free IGF-1 has a circulating half-life of approximately 10-12 minutes. When bound in the IGFBP-3/ALS ternary complex, this extends to roughly 12-15 hours, but the IGF-1 in that complex is biologically inactive until released by proteolytic cleavage of the binding protein. IGF-1 LR3, by contrast, circulates in a free, bioactive state with a functional half-life of approximately 20-30 hours. This is not because LR3 is intrinsically more resistant to degradation (though the N-terminal extension may provide some protection against N-terminal exopeptidases), but primarily because it avoids the rapid clearance that the binding proteins impose on free IGF-1. Renal filtration and receptor-mediated endocytosis become the dominant clearance mechanisms.
Increased potency. Because a far greater proportion of administered LR3 is available in free form to activate the IGF-1R, the effective potency per unit dose is approximately 2-3 times higher than equimolar native IGF-1. Some in vitro assays have reported even greater differences depending on the cell type and the IGFBP milieu in the culture system. In cell culture media supplemented with serum (which contains IGFBPs), LR3 consistently outperforms native IGF-1 because the native peptide gets captured by binding proteins while LR3 does not.
Preserved Receptor Activity
Despite the structural modifications, IGF-1 LR3 retains full agonist activity at the type 1 IGF receptor. The receptor binding site on IGF-1 primarily involves residues in the B and A domains of the protein (the central part of the molecule), which are left intact by the LR3 modifications. The N-terminal extension and position 3 substitution fall outside the critical receptor contact regions, allowing the modified protein to engage and activate IGF-1R with comparable efficacy to the native ligand.
This means that the entire downstream signaling cascade described in the biology section - PI3K/Akt/mTOR activation, FoxO phosphorylation, MAPK/ERK signaling - is triggered by LR3 just as effectively as by native IGF-1. The difference is purely pharmacokinetic: LR3 maintains this receptor activation for a much longer period and at effectively higher concentrations relative to the administered dose.
LR3 also retains modest cross-reactivity with the insulin receptor, as does native IGF-1. At physiological concentrations this is generally not clinically significant, but at the supraphysiological levels achievable with exogenous LR3 administration, the insulin-receptor activation contributes to the hypoglycemic effects discussed in the safety section.
Half-Life Comparison Across IGF-1 Variants
Circulating Half-Life: IGF-1 Variants
Data represents approximate circulating half-lives. Native IGF-1 value reflects the free (unbound) fraction. LR3 value represents the functionally active circulating duration. DES value reflects the ultra-short truncated variant.
The chart above illustrates the dramatic differences in circulating duration across the three main IGF-1 variants. Native IGF-1 (free fraction) is cleared in roughly 12 minutes. IGF-1 DES (Des(1-3) IGF-1), a truncated variant missing the first three N-terminal amino acids, has an even shorter half-life of approximately 20-30 minutes but compensates with up to 10-fold higher receptor binding affinity. IGF-1 LR3, at roughly 1,200 minutes (20 hours), represents a fundamentally different pharmacokinetic class entirely.
The Significance of Continuous vs. Pulsatile Signaling
One of the most underappreciated aspects of IGF-1 LR3's modified pharmacokinetics is the shift from pulsatile to continuous receptor stimulation. In normal physiology, IGF-1 signaling follows a dynamic pattern. Growth hormone is released in pulses (predominantly during sleep), triggering waves of hepatic IGF-1 production. Local muscle IGF-1 (including MGF) is produced transiently in response to mechanical loading. IGFBP proteases are activated in a tissue-specific, context-dependent manner to release bioactive IGF-1 precisely where and when it's needed.
This pulsatile pattern isn't just an artifact of physiology - it's functionally important. Continuous vs. pulsatile stimulation of the same receptor can produce qualitatively different downstream effects. In the GH-IGF-1 axis, pulsatile GH secretion is known to be more effective at stimulating IGF-1 gene transcription than continuous infusion at the same total dose. Similarly, intermittent IGF-1 receptor activation may produce different effects on satellite cell dynamics, protein turnover balance, and receptor sensitivity than the sustained activation that LR3's long half-life creates.
For researchers working with growth hormone releasing peptides like GHRP-2, GHRP-6, or sermorelin, one advantage of these compounds is that they preserve the natural pulsatile pattern of the GH-IGF-1 axis rather than overriding it. LR3, by contrast, creates a pharmacological state that has no natural physiological equivalent: sustained, unregulated, high-level IGF-1 receptor activation throughout the dosing interval.
LR3 in Cell Culture and Biotechnology
It's worth understanding that IGF-1 LR3 was not originally developed for therapeutic use. Its primary commercial application has been as a cell culture supplement in the biopharmaceutical industry. When growing mammalian cells for the production of recombinant proteins (monoclonal antibodies, enzymes, etc.), cells require growth factor support. Native IGF-1 works poorly in this role because serum-containing or chemically defined media also contain IGFBPs that sequester it. LR3's resistance to IGFBP binding makes it far more effective at supporting cell proliferation in culture, typically at concentrations of 50-100 ng/mL.
Companies like GroPep Bioreagents (now Novozymes) and others have supplied LR3 for this purpose for decades. The established commercial production pipeline means that research-grade LR3 is relatively accessible, but the quality, purity, and sterility of products marketed for human use outside regulated pharmaceutical channels varies enormously. This is an important practical consideration addressed in the dosing section.
Structural Variants of IGF-1: A Family of Modifications
IGF-1 LR3 is just one of several engineered IGF-1 variants. Understanding the broader family provides useful context:
| Variant | Modification | Amino Acids | IGFBP Binding | Half-Life | Primary Use |
|---|---|---|---|---|---|
| Native IGF-1 | None (endogenous) | 70 | Full affinity | ~12 min (free) | Endogenous hormone |
| Mecasermin (Increlex) | Recombinant native | 70 | Full affinity | ~5.8 hours | FDA-approved (IGFD) |
| IGF-1 LR3 | N-extension + Glu3Arg | 83 | ~100x reduced | ~20-30 hours | Research / cell culture |
| IGF-1 DES | N-terminal truncation | 67 | Very low | ~20-30 min | Research |
| R3-IGF-1 | Glu3Arg only | 70 | Moderately reduced | Not well characterized | Research only |
Each variant represents a different strategy for modifying the balance between receptor potency, IGFBP regulation, and duration of action. The selection among these for research purposes depends on whether the goal is localized, short-duration receptor activation (IGF-1 DES), sustained systemic exposure (IGF-1 LR3), or preserved physiological regulation (native/mecasermin).
Pharmacodynamic Implications of IGFBP Escape
To fully appreciate the pharmacological implications of LR3's reduced IGFBP binding, consider what the binding protein system actually does in normal physiology. IGFBPs don't just extend IGF-1's half-life and regulate its bioavailability - they direct IGF-1 to specific tissues, create concentration gradients, and modulate receptor interactions in ways that shape the biological response. IGFBP-5, for example, can concentrate IGF-1 at the cell surface by binding to extracellular matrix proteins, effectively creating a local reservoir that releases IGF-1 in proximity to its receptor. IGFBP-3 can translocate to the cell nucleus and directly modulate transcription independent of IGF-1.
When LR3 escapes this entire regulatory layer, several consequences follow:
Loss of tissue selectivity. Normal IGF-1 is preferentially directed to specific tissues by differential IGFBP expression and proteolytic release. IGFBP-5 concentrates IGF-1 in bone and muscle; IGFBP-4 in kidney; IGFBP-2 in brain. LR3, free from binding protein guidance, distributes according to simple pharmacokinetic principles (blood flow, tissue permeability) rather than the sophisticated targeting that binding proteins provide. This means that while native IGF-1 might preferentially act on muscle and bone in the post-exercise state, LR3 activates receptors equally in all accessible tissues, including those where sustained IGF-1 signaling may be harmful.
Loss of temporal regulation. IGFBP proteases are activated in specific physiological contexts to release bioactive IGF-1 precisely when and where it's needed. Pregnancy-associated plasma protein A (PAPP-A) cleaves IGFBP-4 in response to inflammatory signals; matrix metalloproteinases cleave IGFBP-5 during tissue remodeling. LR3 bypasses this temporal control, providing constant receptor activation regardless of whether the tissue "needs" IGF-1 signaling at that moment.
Loss of dose modulation. The binding protein system acts as a buffer, absorbing excess IGF-1 and releasing it gradually. This prevents the sharp peaks and troughs in receptor activation that would otherwise occur with each GH pulse. LR3's receptor activation follows its circulating concentration directly, without buffering. After injection, receptor activation peaks as drug levels peak, then declines as the drug is cleared, creating a pharmacokinetic profile more similar to a direct-acting drug than to the naturally regulated hormone system.
The Role of ALS in the Ternary Complex
The acid-labile subunit (ALS) is a liver-derived protein that forms the 150 kDa ternary complex with IGFBP-3 (or IGFBP-5) and IGF-1. This complex is the primary reservoir of circulating IGF-1, carrying approximately 75-80% of the total. ALS serves two key functions: it prevents the IGF-1/IGFBP binary complex from crossing capillary barriers (keeping IGF-1 in the vascular compartment), and it stabilizes the complex against proteolysis, extending its circulating persistence.
ALS production is itself regulated by GH, creating a feedforward loop: GH stimulates both IGF-1 and ALS production, and the resulting ternary complex serves as a circulating reservoir that smooths out the pulsatile pattern of GH-stimulated IGF-1 secretion. ALS-null mice have dramatically reduced circulating IGF-1 despite normal hepatic production, with the unbound IGF-1 being rapidly cleared by renal filtration.
LR3's inability to form the ternary complex with ALS (because it doesn't bind IGFBP-3 or IGFBP-5 with sufficient affinity) means it is cleared more like free IGF-1 than like IGF-1 in the ternary complex. Yet because it's a larger molecule than free IGF-1 (83 vs. 70 amino acids, with the N-terminal extension potentially reducing renal filtration efficiency), its clearance is somewhat slower than that of truly free native IGF-1. This intermediate clearance rate - slower than free IGF-1 but without the massive prolongation that the ternary complex provides - accounts for LR3's observed 20-30 hour half-life.
Implications for Research Design
For researchers designing experiments with IGF-1 variants, the choice between LR3, DES, and native IGF-1 should be driven by the specific biological question:
- If studying IGF-1R signaling per se (pathway activation, gene expression changes, downstream targets), LR3 is often preferred because it provides sustained, consistent receptor activation without the confounding variable of IGFBP modulation. Its extended half-life simplifies dosing schedules in animal studies.
- If studying physiological IGF-1 signaling (the integrated response including IGFBP regulation, tissue targeting, temporal dynamics), native IGF-1 is more appropriate because it preserves the regulatory architecture that shapes the biological response in vivo.
- If studying localized, acute IGF-1R activation, DES is preferred because its rapid clearance limits exposure to the injection site and prevents the chronic systemic receptor activation that complicates interpretation of prolonged LR3 exposure.
- For cell culture applications, LR3 remains the gold standard because its resistance to IGFBP sequestration provides consistent, predictable receptor activation in serum-containing or chemically defined media.
The peptide research hub provides additional guidance on experimental design considerations for growth factor peptides, including practical tips for reconstitution, storage, and dosing calculations across different model systems.
Muscle Hypertrophy Research
How Does IGF-1 LR3 Affect Muscle Growth?
IGF-1 LR3 promotes skeletal muscle growth through two mechanistically distinct pathways: muscle fiber hypertrophy (increasing the size of existing muscle fibers through enhanced protein synthesis and reduced protein degradation) and satellite cell-mediated hyperplasia (increasing the number of myonuclei available to support larger fiber volumes through activation and fusion of muscle stem cells). The relative contribution of each pathway depends on the dose, duration of exposure, and the physiological state of the muscle tissue.
The evidence base for IGF-1's muscle growth effects is extensive, though it draws primarily from native IGF-1 and transgenic animal studies rather than from clinical trials specifically using the LR3 analog. Because LR3 activates the identical receptor and downstream signaling pathways, the mechanistic findings from native IGF-1 research are directly applicable, with the caveat that LR3's extended duration and higher effective potency may amplify both beneficial and adverse effects.
Preclinical Evidence: Animal Models
The most compelling preclinical evidence comes from several landmark studies that established IGF-1 as a direct regulator of muscle mass independent of the systemic GH-IGF-1 axis:
Localized IGF-1 infusion studies. Adams and McCue (1998) demonstrated that localized infusion of IGF-1 directly into rat hindlimb muscles produced significant hypertrophy of approximately 9% in muscle wet weight within 14 days, without affecting contralateral (non-infused) muscles. This provided definitive evidence for a direct local action of IGF-1 on muscle tissue, independent of systemic effects. The hypertrophy was accompanied by increased protein synthesis rates and satellite cell activation, as measured by BrdU incorporation (Adams GR, McCue SA. Localized infusion of IGF-I results in skeletal muscle hypertrophy in rats. J Appl Physiol. 1998;84(5):1716-1722).
Transgenic muscle-specific IGF-1 overexpression. The landmark study by Musaro, McCullagh, Paul and colleagues published in Nature Genetics in 2001 used a tissue-restricted transgene encoding a locally acting IGF-1 isoform (mIGF-1) expressed specifically in skeletal muscle. The transgenic mice developed normally during embryogenesis, and postnatal increases in muscle mass and strength occurred without the pathological organ enlargement or metabolic disturbances seen in models with systemic IGF-1 excess. Most remarkably, the aged transgenic mice (24+ months) maintained muscle mass and regenerative capacity comparable to young wild-type animals, while age-matched controls showed the expected sarcopenic decline (Musaro A, McCullagh K, Paul A, et al. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nature Genetics. 2001;27(2):195-200. DOI: 10.1038/84839).
This study was particularly significant because it demonstrated that locally elevated IGF-1 could preserve muscle function during aging without the cancer risk, organ enlargement, and metabolic disruption associated with systemic IGF-1 elevation. The distinction between local and systemic effects is a recurring theme in this field.
Long R3 IGF-1 systemic infusion in guinea pigs. Lok and colleagues (1996) infused Long R3 IGF-1 (the LR3 variant) systemically in guinea pigs at 1.5 mg/kg/day for 7 days. The treatment stimulated significant organ growth, including increased weights of spleen, kidneys, and adrenals. However, it simultaneously reduced circulating levels of endogenous IGF-1, IGF-2, and several IGFBPs, demonstrating that exogenous LR3 suppresses the endogenous IGF system through feedback inhibition (Lok F, Owens JA, Mundy L, et al. Long R3 insulin-like growth factor-I (IGF-I) infusion stimulates organ growth but reduces plasma IGF-I, IGF-II and IGF binding protein concentrations in the guinea pig. J Endocrinol. 1996;148(1):77-87. PMID: 7561636).
The Satellite Cell Connection
Satellite cells are muscle-specific stem cells that reside between the basal lamina and the sarcolemma of mature muscle fibers. In adult muscle, they exist primarily in a quiescent state, entering the cell cycle only in response to specific activation signals, including mechanical damage, exercise-induced microtrauma, and growth factor stimulation. IGF-1 is one of the most potent activators of satellite cell proliferation and differentiation.
Rosenblatt and Parry (1992) elegantly demonstrated the contribution of satellite cells to IGF-1-induced muscle growth using gamma radiation to selectively ablate proliferating satellite cells. In rats receiving IGF-1 infusion, gamma irradiation of the treated muscle prevented approximately 50% of the IGF-1-induced hypertrophy. This finding established that satellite cell-mediated nuclear addition accounts for roughly half of IGF-1's growth effect, with the remainder attributable to increased protein synthesis per existing myonucleus (Rosenblatt JD, Parry DJ. Gamma irradiation prevents compensatory hypertrophy of overloaded mouse extensor digitorum longus muscle. J Appl Physiol. 1992;73(6):2538-2543).
In human satellite cell cultures, IGF-1 treatment extended the proliferative lifespan by approximately 4.3% (from 46 to 48 mean population doublings) and promoted myogenic differentiation, confirming that human muscle stem cells are responsive to IGF-1 signaling. Subsequent work demonstrated that IGF-1-induced hypertrophy in human myotube cultures involved both increased protein synthesis per nucleus and recruitment of additional nuclei from the reserve cell pool (Jacquemin V, Furling D, Bigot A, et al. IGF-1 induces human myotube hypertrophy by increasing cell recruitment. Exp Cell Res. 2004;299(1):148-158).
Mechanistic Pathways in Detail
The molecular mechanisms through which IGF-1 (and by extension, IGF-1 LR3) drives muscle hypertrophy are now well characterized:
Protein synthesis acceleration. The PI3K/Akt/mTOR axis, as described in the biology section, directly increases the rate of muscle protein synthesis. In rodent models, IGF-1 administration increases mixed muscle protein synthesis rates by 20-40% within hours of exposure. The mTORC1-mediated phosphorylation of p70S6K and 4E-BP1 increases both ribosomal biogenesis (making more protein-manufacturing machinery) and translation initiation efficiency (making each ribosome work faster).
Proteolysis inhibition. The Akt-FoxO axis is equally important for net protein accretion. Phosphorylated (inactive) FoxO transcription factors cannot enter the nucleus to drive expression of the atrogenes MuRF1 and MAFbx/atrogin-1. These E3 ubiquitin ligases are the primary executors of muscle protein breakdown through the ubiquitin-proteasome system. By simultaneously stimulating synthesis and suppressing degradation, IGF-1/Akt shifts the protein balance strongly toward net accretion (Stitt TN, Drujan D, Clarke BA, et al. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol Cell. 2004;14(3):395-403).
Myoblast proliferation and fusion. Through the MAPK/ERK pathway, IGF-1 drives satellite cell entry into the cell cycle, proliferation, and subsequent differentiation into myocytes that fuse with existing fibers (contributing new nuclei) or, in some circumstances, fuse with each other to form new fibers (hyperplasia). The "myonuclear domain" hypothesis suggests that each myonucleus can support only a finite volume of cytoplasm, so substantial hypertrophy beyond a certain point requires nuclear addition from satellite cells.
Anti-apoptotic protection. During periods of muscle stress (eccentric exercise-induced damage, disuse, inflammation), a significant amount of muscle protein loss results from myonuclear apoptosis. IGF-1's activation of the PI3K/Akt survival pathway protects myonuclei from apoptotic death, preserving the fiber's nuclear-to-cytoplasmic ratio and maintaining its capacity for protein production.
Human Studies with Recombinant IGF-1
Direct human trials with IGF-1 for muscle growth are limited, but the clinical experience with mecasermin (recombinant human IGF-1) provides relevant data. In the registration trials for severe primary IGF-1 deficiency in children, subcutaneous mecasermin at 0.12 mg/kg twice daily increased height velocity from a mean of 2.8 cm/year at baseline to 8.0 cm/year during the first year, with sustained effects over 8+ years of treatment (Chernausek SD, Backeljauw PF, Frane J, et al. Long-term treatment with recombinant IGF-I in children with severe IGF-I deficiency due to growth hormone insensitivity. J Clin Endocrinol Metab. 2007;92(3):902-910).
In adult populations, the evidence for IGF-1's muscle effects comes primarily from observational studies and GH replacement trials (since GH works largely through increasing endogenous IGF-1). Meta-analyses of GH therapy in older adults show modest increases in lean body mass (approximately 2 kg) but inconsistent effects on muscle strength and functional outcomes, suggesting that systemic IGF-1 elevation alone may not be sufficient to produce clinically meaningful improvements in muscle performance without concurrent resistance training.
This underscores an important practical point: IGF-1 (or LR3) is unlikely to produce significant muscle growth in the absence of appropriate mechanical loading and nutritional support. The peptide amplifies the anabolic response to training; it doesn't replace it. For those interested in how growth hormone secretagogues compare in this regard, our coverage of MK-677 (Ibutamoren) examines the evidence for GH-axis stimulation and body composition changes in detail.
Additional context from GH replacement trials in adults highlights the importance of combined stimuli. In elderly subjects receiving GH replacement (which increases IGF-1), lean mass increased by approximately 2 kg over several months, but without concurrent resistance training, there was no meaningful improvement in strength, power, or physical function. When GH therapy was combined with resistance training, the functional outcomes improved significantly, suggesting that IGF-1 elevation provides the permissive biochemical environment for muscle adaptation but requires the mechanical stimulus to direct that environment toward functional improvement.
This finding is consistent with the molecular biology: mTOR activation in response to both IGF-1 signaling and mechanical loading converges on the same downstream effectors (p70S6K, 4E-BP1), but mechanical loading also activates unique mechanosensitive pathways (phospholipase D, ERK1/2 in a contraction-dependent manner) that contribute to the specificity of the hypertrophic response. Without the mechanical input, IGF-1-driven protein synthesis may result in protein accretion without the organized sarcomeric assembly that produces functional muscle.
IGF-1 and Muscle Regeneration in Aging
One of the most compelling potential applications of IGF-1 signaling relates to age-related muscle decline. Sarcopenia - the progressive loss of skeletal muscle mass and function with aging - affects an estimated 10-16% of adults over age 60 and contributes significantly to frailty, falls, disability, and mortality. The age-related decline in both systemic and local muscle IGF-1 production is considered a contributing factor.
The Musaro et al. (2001) transgenic mouse study mentioned earlier demonstrated that muscle-specific IGF-1 overexpression could maintain youthful muscle mass and regenerative capacity into old age. The aged transgenic mice expressed GATA-2, a transcription factor normally absent in skeletal muscle, which marked hypertrophic myocytes that had escaped the typical sarcopenic decline. These muscle fibers retained the satellite cell proliferative response to injury that is normally diminished in aged animals.
A related study using the same mIGF-1 transgenic model showed that localized IGF-1 expression attenuated disuse atrophy during hindlimb unloading, suggesting protection against both age-related and disuse-mediated muscle wasting (Musaro A, Giacinti C, Borsellino G, et al. Paracrine effects of IGF-1 overexpression on the functional decline due to skeletal muscle disuse. PLoS One. 2013;8(5):e65167).
These findings generate enthusiasm but also require careful interpretation. The transgenic models achieve localized, constitutive, moderate-level IGF-1 elevation restricted to muscle tissue. This is pharmacologically quite different from injecting IGF-1 LR3 systemically, which exposes every tissue in the body to elevated IGF-1 receptor activation. The anti-sarcopenia potential may be real, but the systemic approach carries risks that the transgenic approach avoids.
Readers interested in other approaches to age-related tissue decline may find our reports on Epithalon and Humanin relevant, as these compounds target different aspects of the aging process through complementary mechanisms.
What the Evidence Actually Supports
Taking the full body of evidence together, several conclusions are well-supported:
- IGF-1 receptor activation directly promotes muscle protein synthesis and inhibits proteolysis through clearly defined molecular pathways.
- IGF-1 activates satellite cells, and satellite cell-mediated nuclear addition accounts for approximately half of IGF-1-induced hypertrophy.
- Localized IGF-1 elevation in muscle tissue preserves muscle mass and regenerative capacity during aging in animal models.
- Systemic IGF-1 elevation produces muscle growth but also causes organ enlargement, metabolic disturbance, and potentially increased cancer risk.
- IGF-1 LR3 is expected to produce qualitatively similar but quantitatively amplified muscle effects compared to native IGF-1, given its higher effective potency and longer duration of action.
What the evidence does not support is a clear dose-response relationship for LR3 in human muscle growth, an established safety margin for chronic use, or evidence that LR3 produces superior functional outcomes (strength, power, endurance) compared to other anabolic interventions like resistance training plus adequate protein intake. These gaps represent the critical unknowns that anyone considering this compound should acknowledge.
The Myonuclear Domain Theory and IGF-1
The myonuclear domain theory posits that each myonucleus controls the gene expression and protein production for a finite volume of cytoplasm surrounding it. In human muscle fibers, each myonucleus is estimated to control a domain of approximately 2,000-2,500 square micrometers of cross-sectional area. When a fiber hypertrophies beyond the capacity of its existing myonuclei, new nuclei must be recruited from satellite cells to maintain the nuclear-to-cytoplasmic ratio. Without nuclear addition, the domain becomes "overstretched" and the fiber cannot maintain its larger size.
This theory has direct implications for IGF-1 LR3's mechanism of action. Short-term hypertrophy (the first few weeks of anabolic stimulus) can occur through increased protein synthesis alone, without satellite cell contribution, as existing myonuclei simply increase their biosynthetic output. But sustained, substantial hypertrophy (beyond approximately 15-26% fiber cross-sectional area increase in humans) appears to require satellite cell-mediated nuclear addition.
Petrella and colleagues (2008) demonstrated this in a landmark human study examining the relationship between satellite cell number and muscle fiber hypertrophy during 16 weeks of resistance training. Using cluster analysis, they identified "extreme responders" (mean fiber hypertrophy of 54%) who had significantly higher satellite cell content and greater myonuclear addition compared to "modest responders" (mean 28% hypertrophy) and "non-responders" (no significant hypertrophy). The extreme responders showed 23% more satellite cells at baseline and 48% greater expansion of the satellite cell pool during training (Petrella JK, Kim JS, Mayhew DL, Cross JM, Bamman MM. Potent myofiber hypertrophy during resistance training in humans is associated with satellite cell-mediated myonuclear addition. J Appl Physiol. 2008;104(6):1736-1742. DOI: 10.1152/japplphysiol.01215.2007).
IGF-1's dual action on both protein synthesis (through PI3K/Akt/mTOR) and satellite cell activation (through MAPK/ERK and autocrine IGF-1 signaling in satellite cells) positions it as a potentially effective promoter of both phases of hypertrophy. The LR3 variant, with its extended duration and enhanced bioavailability, may provide more sustained satellite cell stimulation than native IGF-1, though this has not been directly tested in human subjects.
IGF-1 and Muscle Fiber Type Specificity
Human skeletal muscle contains a spectrum of fiber types ranging from slow-twitch oxidative (Type I) to fast-twitch glycolytic (Type IIx/IIb), with intermediate hybrid fibers (Type IIa). These fiber types differ in their contractile properties, metabolic characteristics, and response to various growth stimuli. IGF-1 signaling appears to preferentially promote hypertrophy of Type II (fast-twitch) fibers, which have higher IGF-1R expression density and greater mTOR pathway responsiveness than Type I fibers.
This fiber type preference is clinically relevant because Type II fibers are the fibers most affected by age-related sarcopenia. Adults lose Type II fiber cross-sectional area more rapidly than Type I with advancing age, contributing to the disproportionate loss of strength and power relative to total muscle mass in older adults. The preferential effect of IGF-1 on Type II fibers suggests that it might be particularly effective at counteracting this specific aspect of aging, though translating this from mechanism to clinical outcome requires evidence that doesn't yet exist for LR3.
Interestingly, the transgenic mice in the Musaro et al. study expressed the mIGF-1 transgene under a muscle-specific promoter active in all fiber types, yet the protective effect against age-related atrophy appeared most pronounced in fast-twitch muscles. This suggests that even with uniform IGF-1 exposure, the downstream response is modulated by fiber-type-specific factors in the signal transduction machinery or gene expression program.
Hyperplasia vs. Hypertrophy: Distinguishing the Growth Mechanisms
One of the most debated topics in muscle biology is whether skeletal muscle in adult humans is capable of true hyperplasia (formation of entirely new muscle fibers) or whether all post-natal muscle growth occurs through hypertrophy (enlargement of existing fibers with satellite cell-mediated nuclear addition). This distinction matters for understanding IGF-1 LR3's potential effects because the compound's proponents often claim it can promote muscle fiber hyperplasia, a property that would be unique among currently available anabolic agents.
In animal models, the evidence for IGF-1-driven hyperplasia is mixed. In avian muscle, IGF-1 can clearly promote the formation of new muscle fibers from satellite cells. In rodent models, there is some evidence for fiber splitting and new fiber formation following IGF-1 treatment, particularly in young growing animals or in the context of muscle damage. However, distinguishing true de novo fiber formation from regenerative repair of damaged fibers is technically challenging, and some apparent "hyperplasia" may actually represent fiber regeneration or fiber splitting events rather than net new fiber creation.
In adult human muscle, the evidence for hyperplasia is extremely limited. Most studies of resistance training-induced muscle growth show increases in fiber cross-sectional area (hypertrophy) without increases in total fiber number. The human muscle fiber number appears to be largely established during prenatal development and early infancy, with relatively little capacity for new fiber formation in adults under physiological conditions.
However, there is a theoretical basis for IGF-1 to promote some degree of hyperplasia even in adult muscle. When satellite cells are activated and proliferate in response to IGF-1, most myoblasts fuse with existing fibers (contributing to hypertrophy). But a subset of activated satellite cells can fuse with each other to form new, small myotubes that may mature into independent fibers. Whether this occurs to any meaningful degree with systemic LR3 administration in humans is unknown, and claims of LR3-driven hyperplasia should be viewed as unsubstantiated by current evidence.
The distinction has practical implications beyond academic interest. If LR3 primarily produces hypertrophy, its effects are mechanistically similar (though potentially greater in magnitude) to those achievable through resistance training plus adequate nutrition. If it produces true hyperplasia - increasing the total number of muscle fibers available for training-induced growth - the effects would be qualitatively different and potentially more permanent, as additional fibers would persist even after LR3 use is discontinued. The absence of definitive human data on this question is a significant gap in the literature.
IGF-1 and Connective Tissue: Tendons, Ligaments, and Joints
While the focus on muscle growth dominates discussions of IGF-1 LR3, the compound's effects on connective tissue deserve attention because they have practical implications for musculoskeletal health. IGF-1 promotes collagen synthesis by fibroblasts, stimulates proteoglycan production in cartilage, and enhances extracellular matrix formation in tendons and ligaments.
In theory, IGF-1's connective tissue effects could be beneficial by strengthening the tendons and ligaments that must support larger, stronger muscles. Rapid muscle hypertrophy without corresponding connective tissue adaptation is a recognized risk factor for tendon and ligament injuries, and the slower remodeling rate of connective tissue compared to muscle means that structural tissues often lag behind muscle growth during periods of aggressive anabolic stimulation.
However, the evidence for IGF-1's effects on connective tissue strength in vivo is limited. In vitro, IGF-1 stimulates tendon fibroblast proliferation and collagen synthesis. In animal models, local IGF-1 application has shown some benefits for tendon healing. But systemic IGF-1 elevation may have different effects than local application, and the quality and mechanical properties of IGF-1-stimulated connective tissue (not just its quantity) determine functional benefit.
In acromegaly patients, who have chronically elevated IGF-1, joint problems are common: arthropathy (joint disease) affects 50-70% of patients, with cartilage hypertrophy initially followed by progressive joint degeneration. This pattern - initial cartilage growth followed by deterioration - suggests that while short-term IGF-1 elevation may support connective tissue, chronic excess can be destructive. Peptides like BPC-157 and TB-500 offer connective tissue support through mechanisms that don't carry these same risks and may be more appropriate for musculoskeletal health optimization.
The Exercise-IGF-1 Interaction
A critical practical question for anyone considering IGF-1 LR3 is how exogenous IGF-1 interacts with exercise-induced anabolic signaling. Resistance exercise independently activates the mTOR pathway through mechanosensitive signaling (via phospholipase D, diacylglycerol, and phosphatidic acid) and through exercise-induced increases in local IGF-1 (particularly MGF) production. Does adding exogenous LR3 to an already-activated mTOR pathway provide additive benefit, or are the pathways already maximally stimulated by exercise alone?
The available evidence suggests that the answer depends on the training status and nutritional state of the individual. In untrained individuals performing their first bouts of resistance exercise, endogenous mTOR activation is strong and may be near-maximal, leaving limited room for exogenous IGF-1 to add further stimulus. In highly trained individuals who have accumulated years of training adaptation, the acute mTOR response to a given exercise bout is attenuated (a phenomenon called "dampened signaling"), potentially creating more headroom for exogenous IGF-1 to enhance the response.
Nutritional status also matters critically. Protein intake, particularly the leucine content of post-exercise meals, is one of the most potent natural activators of mTOR. A post-exercise meal containing 20-40 grams of high-quality protein with 2-3 grams of leucine produces strong mTOR activation that may overlap substantially with the signaling produced by exogenous IGF-1. This raises the practical question of whether IGF-1 LR3's anabolic effects are additive to optimal training and nutrition, or whether they primarily benefit individuals with suboptimal nutritional or training stimuli.
For individuals interested in optimizing the exercise-IGF-1 interaction without the risks of exogenous IGF-1, strategies that enhance endogenous IGF-1 production deserve consideration. High-intensity resistance training, adequate sleep (supporting GH pulsatility), sufficient protein intake, and avoidance of chronic caloric restriction all support endogenous IGF-1 levels. GH secretagogues like Hexarelin or GHRP-2 can further amplify endogenous GH and IGF-1 within the physiological regulatory framework.
IGF-1 in Muscle Disease and Wasting Conditions
Beyond normal hypertrophy, IGF-1 has been investigated in several muscle wasting conditions where the therapeutic rationale is particularly compelling:
Spinal and Bulbar Muscular Atrophy (SBMA). Palazzolo and colleagues (2009) demonstrated that muscle-specific IGF-1 overexpression in a transgenic mouse model of SBMA (Kennedy disease) attenuated disease progression, reduced toxic protein aggregation, and improved motor function. The IGF-1 effect was mediated through activation of the Akt pathway, which phosphorylated and cleared the mutant androgen receptor protein. This study, published in Neuron, suggested that localized IGF-1 might address the primary disease mechanism rather than merely compensating for muscle loss.
Cancer-related cachexia. Muscle wasting in cancer patients (cachexia) involves activation of the ubiquitin-proteasome system through increased MuRF1 and MAFbx expression - precisely the pathway that IGF-1/Akt signaling suppresses. Theoretically, IGF-1 could counteract cachexia-associated muscle loss. However, the cancer risk implications of administering a growth factor to cancer patients create an obvious contraindication that limits clinical investigation.
Heart failure-associated muscle wasting. Schulze and colleagues demonstrated that muscle-specific IGF-1 overexpression inhibited ubiquitin-mediated muscle atrophy in a chronic heart failure model in mice. The transgenic mice showed preserved muscle mass and reduced expression of atrogenes despite cardiac dysfunction (published in Circulation Research, 2005). This provides preclinical support for IGF-1-based interventions in cardiac cachexia, though systemic administration carries its own cardiovascular risks.
Disuse atrophy. Prolonged bed rest, limb immobilization, and spaceflight all produce rapid muscle atrophy. The Musaro lab's work with mIGF-1 transgenic mice showed significant protection against hindlimb unloading-induced atrophy, suggesting that IGF-1 signaling could mitigate disuse-related muscle loss. This has implications for post-surgical recovery, critical care patients, and astronauts on long-duration spaceflight.
Duchenne Muscular Dystrophy (DMD). DMD is a progressive muscle wasting disease caused by mutations in the dystrophin gene. While IGF-1 cannot replace the missing dystrophin protein, its anti-atrophic and satellite-cell-activating properties have generated interest as a potential adjunctive therapy. Preclinical studies in the mdx mouse model of DMD showed that IGF-1 overexpression preserved muscle function and reduced fibrosis. However, the inflammatory and fibrotic environment in DMD muscle may alter IGF-1 signaling dynamics compared to healthy muscle, and the oncogenic concerns limit enthusiasm for systemic IGF-1 approaches in young patients with decades of potential exposure ahead of them.
Sarcopenic Obesity. The combination of excess body fat and reduced muscle mass (sarcopenic obesity) is increasingly recognized as a high-risk metabolic phenotype associated with greater disability, metabolic dysfunction, and mortality than either condition alone. IGF-1's dual effects on muscle (anabolic) and metabolism (insulin-like) make its risk-benefit profile in this population particularly complex. The metabolic effects of systemic IGF-1 elevation in the setting of existing insulin resistance could worsen metabolic dysfunction, even while improving muscle mass. This population might benefit more from approaches that address both components: GLP-1 receptor agonists for the obesity component, combined with resistance training and potentially GH secretagogues for the muscle component.
These disease-specific applications highlight the therapeutic potential of targeted IGF-1 signaling in muscle. However, they also underscore the gap between localized, transgenic IGF-1 expression (which has been repeatedly shown to be effective and relatively safe in animal models) and systemic administration of LR3 (which has not been tested in these clinical contexts). The challenge remains to achieve muscle-targeted IGF-1 signaling without the systemic exposure that creates risk for other tissues. Gene therapy approaches, tissue-specific delivery systems, and engineered IGF-1 variants with improved tissue targeting represent active areas of research that may eventually bridge this gap.
Comparison to Other Growth Factors
How Does IGF-1 LR3 Compare to Other Growth-Promoting Compounds?
IGF-1 LR3 exists within a broader family of compounds that influence the growth hormone-IGF-1 axis and anabolic signaling. Each compound in this family has distinct mechanisms, pharmacokinetic profiles, regulatory status, and risk-benefit ratios. Understanding where LR3 fits relative to alternatives is essential for informed decision-making, whether in research design or clinical practice.
IGF-1 LR3 vs. IGF-1 DES (Des(1-3) IGF-1)
IGF-1 DES takes the opposite engineering approach to LR3. Where LR3 adds amino acids to the N-terminus, DES removes the first three, creating a 67-amino-acid truncated peptide. This deletion eliminates a key IGFBP binding region, reducing binding protein affinity similarly to LR3, but the truncation also increases receptor binding affinity by up to 10-fold compared to native IGF-1.
The critical difference is pharmacokinetic. IGF-1 DES has a very short half-life of approximately 20-30 minutes, making it essentially a "pulse" of intense, localized IGF-1 receptor activation. When injected intramuscularly at a specific site, the high receptor affinity and rapid local action may preferentially activate satellite cells in the injected muscle before the peptide is cleared systemically. This localized action profile has made DES popular among researchers interested in site-specific muscle effects.
LR3, by contrast, is a systemic agent. Its 20-30 hour half-life means it distributes throughout the body and activates IGF-1 receptors in all tissues for an extended period. This makes it less suitable for targeted local effects but more effective for whole-body anabolic signaling.
| Parameter | IGF-1 LR3 | IGF-1 DES |
|---|---|---|
| Amino acids | 83 | 67 |
| Receptor binding affinity | ~Equal to native IGF-1 | ~10x higher than native |
| IGFBP binding | ~100x reduced | Very low / negligible |
| Half-life | 20-30 hours | 20-30 minutes |
| Systemic exposure | High (prolonged) | Minimal (rapid clearance) |
| Site-specific effects | Limited (distributes widely) | Higher (acts locally before clearance) |
| Hypoglycemia risk | Higher (sustained exposure) | Lower (brief exposure) |
| Cancer risk concern | Higher (chronic receptor activation) | Lower (transient activation) |
IGF-1 LR3 vs. Growth Hormone Secretagogues
Growth hormone secretagogues (GHS) represent a fundamentally different approach to enhancing the GH-IGF-1 axis. Rather than directly supplying the downstream effector (IGF-1), these compounds stimulate endogenous GH production, which then increases IGF-1 through normal physiological pathways. This preserves feedback regulation and pulsatile signaling patterns.
CJC-1295/Ipamorelin is a widely used combination that pairs a GHRH analog (CJC-1295) with a ghrelin receptor agonist (Ipamorelin). CJC-1295 with DAC (Drug Affinity Complex) extends the half-life to approximately 6-8 days, producing sustained GH elevation. The resulting IGF-1 increase is typically 50-100% above baseline, well within the physiological range. This approach raises IGF-1 gradually and maintains regulatory feedback, in stark contrast to the pharmacological "override" that exogenous LR3 represents.
MK-677 (Ibutamoren) is an oral ghrelin receptor agonist that increases GH secretion and consequently elevates IGF-1 levels. In clinical trials, MK-677 at 25 mg daily increased IGF-1 to youthful levels in elderly subjects and produced modest increases in lean body mass. The oral route and preservation of pulsatile GH patterns make it more convenient and physiologically compatible than direct IGF-1 injection, though the IGF-1 elevation is considerably more modest than what LR3 administration achieves.
Sermorelin and Tesamorelin are GHRH analogs that stimulate pituitary GH release. Tesamorelin is FDA-approved for lipodystrophy in HIV patients and has demonstrated reductions in visceral adipose tissue. These compounds work entirely within the physiological regulatory framework, making their safety profile better characterized than direct IGF-1 analogs.
The key tradeoff is clear: GH secretagogues produce more modest, physiologically regulated IGF-1 elevation with better established safety profiles but slower, less dramatic anabolic effects. IGF-1 LR3 produces more intense, unregulated receptor activation with potentially greater anabolic effects but substantially higher risk. For most clinical scenarios, the GH secretagogue approach offers a more favorable risk-benefit ratio.
It's worth examining the secretagogue approach in more detail because it illustrates how preserving physiological regulation can maintain many of IGF-1's benefits while avoiding its most dangerous effects. When CJC-1295/Ipamorelin stimulates GH release from the pituitary, several protective mechanisms remain intact: somatostatin feedback prevents excessive GH secretion; GH-induced IGF-1 production occurs primarily in the liver, where it's immediately captured by IGFBPs; the resulting IGF-1 elevation is modest (typically 50-100% above baseline) and remains within the physiological range; and the natural pulsatile pattern of GH release is preserved, which appears to be important for the metabolic and tissue-specific effects of the axis.
By contrast, injecting LR3 directly introduces IGF-1R agonist activity at levels and durations that no endogenous regulatory mechanism can moderate. There is no IGFBP buffering, no somatostatin brake, no feedback suppression of the exogenous compound itself. The only limit on receptor activation is the administered dose and the compound's clearance kinetics. This creates a fundamentally different pharmacological situation, one that may produce faster and more dramatic anabolic effects but at the cost of the safety margins that physiological regulation provides.
Additional GH secretagogues worth considering in this comparison include GHRP-2, which is among the most potent synthetic GH secretagogues; GHRP-6, which also stimulates appetite through ghrelin receptor activation; and Hexarelin, which produces some of the highest GH release per dose but also carries a higher risk of prolactin and cortisol elevation. Each has a somewhat different profile within the secretagogue class, but all share the fundamental advantage of working within the body's regulatory framework rather than overriding it.
IGF-1 LR3 vs. Mecasermin (Increlex)
Mecasermin is recombinant native human IGF-1, structurally identical to the endogenous hormone. It is the only FDA-approved IGF-1 product, indicated specifically for growth failure in pediatric patients with severe primary IGF-1 deficiency (SPIGFD) or GH gene deletion with neutralizing GH antibodies.
In its key multicenter trial of 76 children with severe IGFD, subcutaneous mecasermin at 0.12 mg/kg twice daily increased height velocity from 2.8 cm/year to 8.0 cm/year in the first year, with growth effects sustained over 8 years of follow-up. The safety data from this long-term clinical program is the most comprehensive dataset available for any IGF-1 compound.
Compared to LR3, mecasermin has three key differences: it is subject to normal IGFBP regulation (limiting its effective potency and duration); it requires twice-daily dosing due to its shorter active period; and it has an established safety profile from years of clinical use. The most common adverse effect is hypoglycemia, reported in 42% of treated subjects, with 5 cases of severe hypoglycemia requiring assistance among 71 subjects in the long-term safety analysis. Other notable effects include lymphoid tissue hyperplasia (particularly tonsillar and adenoidal), lipohypertrophy at injection sites, and coarsening of facial features with long-term use.
For researchers and clinicians, mecasermin provides a useful reference point for predicting LR3's effects. If native IGF-1 at FDA-approved doses in a supervised clinical setting produces hypoglycemia in 42% of patients and requires careful meal timing to manage, LR3's enhanced potency and extended duration can reasonably be expected to amplify this risk, particularly in unsupervised settings.
IGF-1 LR3 vs. Myostatin Inhibitors
Myostatin (GDF-8) is a member of the TGF-beta superfamily that acts as a negative regulator of muscle mass. While no myostatin inhibitors are currently approved for clinical use, the concept provides an instructive comparison with IGF-1-based approaches. Myostatin inhibition promotes muscle growth by removing a natural brake on muscle size, whereas IGF-1 promotes growth by stepping on the anabolic accelerator. The two approaches are mechanistically complementary: myostatin inhibition relieves the restriction on how large a muscle can grow, while IGF-1 provides the biosynthetic drive to actually build larger fibers.
Theoretically, this complementary mechanism suggests that combining myostatin inhibition with IGF-1 signaling could produce greater muscle growth than either approach alone. Preclinical data supports this hypothesis: studies in mice with both myostatin knockout and IGF-1 overexpression show greater muscle mass than either genetic modification alone. However, the practical implementation of this combination in humans is far from the evidence base.
From a safety perspective, myostatin inhibition has an important theoretical advantage over IGF-1 elevation: myostatin's expression is largely muscle-specific, so inhibiting it primarily affects skeletal muscle without the broad mitogenic stimulation that IGF-1R activation produces in all tissues. The cancer risk associated with myostatin inhibition appears to be minimal based on animal models and the limited human data from clinical trials of anti-myostatin antibodies (some of which reached Phase II for muscular dystrophy).
IGF-1 LR3 in the Context of GLP-1 Receptor Agonists
The recent explosion of interest in GLP-1 receptor agonists (semaglutide, tirzepatide, liraglutide) for weight loss has created an interesting clinical context for compounds like IGF-1 LR3. GLP-1 agonists produce substantial weight loss (15-25% of body weight with newer agents), but a significant portion of the weight lost is lean tissue, including skeletal muscle. Some studies report that 25-40% of total weight loss during GLP-1 therapy is fat-free mass, raising concerns about sarcopenia, especially in older patients.
This has generated interest in combining GLP-1 agonists with anabolic agents that could preserve or rebuild lean mass during the weight loss phase. IGF-1 LR3 is one candidate for this role, though the combination raises several concerns: the metabolic interaction between IGF-1's glucose-lowering effects and GLP-1's insulin-stimulating and glucose-lowering effects could create complex hypoglycemic dynamics; the appetite-suppressing effects of GLP-1 agonists could make it difficult to consume the post-injection meals required for safe IGF-1 LR3 use; and the long-term metabolic consequences of combining these pharmacologically distinct agents are entirely unstudied.
More conservative approaches to lean mass preservation during GLP-1 therapy include resistance training (the most evidence-based approach), adequate protein intake (1.0-1.2 g/kg target body weight), and potentially GH secretagogues like CJC-1295/Ipamorelin that provide modest IGF-1 elevation within the physiological regulatory framework. The GLP-1 research hub covers the latest evidence on body composition management during incretin therapy. Newer compounds like retatrutide, a triple agonist (GIP/GLP-1/glucagon), are also being investigated, and the retatrutide hub provides in-depth coverage.
IGF-1 LR3 vs. Growth Hormone (Somatropin)
Recombinant human GH (somatropin) has been used clinically for decades, providing the largest safety and efficacy database in this therapeutic class. GH works upstream of IGF-1, and its effects are mediated through both IGF-1-dependent and IGF-1-independent pathways. GH has direct effects on adipocytes (promoting lipolysis), hepatocytes (stimulating IGF-1 production), and other tissues that are not replicated by IGF-1 alone.
Key differences relevant to the comparison:
- GH promotes fat oxidation and reduces visceral adipose tissue; IGF-1 may increase fat deposition due to its insulin-like metabolic effects.
- GH preserves pulsatile, regulated IGF-1 production; exogenous LR3 bypasses this regulation entirely.
- GH has decades of post-market safety surveillance data; LR3 has essentially none in a clinical context.
- GH is FDA-approved for multiple indications including adult GH deficiency, Turner syndrome, short bowel syndrome, and others; LR3 has no regulatory approval.
IGF-1 LR3 vs. Other Anabolic Peptides
Several other peptides in the peptide research space influence muscle growth or tissue repair through distinct mechanisms:
BPC-157 is a gastric pentadecapeptide that promotes tissue healing, angiogenesis, and tendon/ligament repair. Unlike IGF-1, BPC-157 does not directly stimulate muscle protein synthesis or satellite cell proliferation. Its mechanism involves upregulation of growth factor receptors (including the GH receptor), nitric oxide production, and anti-inflammatory signaling. BPC-157/TB-500 blends combine complementary healing pathways. These compounds address tissue repair rather than growth per se, making them complementary rather than competitive with IGF-1.
MOTS-c is a mitochondria-derived peptide that enhances metabolic function, insulin sensitivity, and exercise capacity. It operates through AMPK activation, a pathway that is actually antagonistic to the mTOR-mediated protein synthesis that IGF-1 promotes. MOTS-c is of interest for metabolic health and exercise performance rather than direct anabolic effects.
AOD-9604 and Fragment 176-191 are GH fragments that retain the lipolytic (fat-burning) properties of GH without the growth-promoting effects. They don't significantly increase IGF-1 and are not expected to produce muscle hypertrophy, positioning them as body composition agents focused on fat reduction rather than muscle building.
5-Amino-1MQ is a small molecule NNMT inhibitor that enhances cellular energy metabolism and may support lean body mass indirectly through improved metabolic efficiency. It represents an entirely different mechanistic approach to body composition than IGF-1 pathway activation.
IGF-1 LR3 vs. Anabolic Steroids
While anabolic steroids and IGF-1 LR3 are both used for muscle growth purposes, they operate through fundamentally different mechanisms and have almost entirely non-overlapping pharmacology. Anabolic-androgenic steroids (AAS) work through androgen receptor activation, which directly upregulates muscle protein synthesis genes, increases satellite cell number and myonuclear density, inhibits cortisol-mediated catabolism, and promotes nitrogen retention. Their primary signaling pathway (androgen receptor to DNA transcription) is distinct from the IGF-1R/PI3K/Akt/mTOR cascade, though there is significant cross-talk: androgens increase local muscle IGF-1 production, and IGF-1 may mediate some of the anabolic effects of testosterone.
Key differences relevant to the comparison include the following. Anabolic steroids have decades of clinical use and safety data, well-characterized dose-response relationships, and established protocols for managing side effects and post-cycle recovery (though they carry their own serious risks including cardiovascular toxicity, hepatotoxicity, and reproductive system suppression). IGF-1 LR3 lacks this evidence base entirely.
The mechanisms are complementary rather than redundant. Testosterone increases myonuclear number through satellite cell activation (androgen-receptor-dependent), while IGF-1 independently activates satellite cells through the MAPK/ERK pathway and drives protein synthesis through mTOR. In theory, combining the two pathways could produce additive or complementary effects on muscle growth, but this combination also creates additive risk profiles. Some researchers in the bodybuilding community report using both compound classes together, but this represents an uncharted pharmacological territory with no safety data whatsoever.
The cancer risk profiles also differ significantly. Androgen excess is associated primarily with prostate cancer risk (through androgen-receptor-dependent pathways in prostatic epithelium). IGF-1 excess is associated with breast, prostate, and colorectal cancer risk through the PI3K/Akt/mTOR pathway. Combined use theoretically creates additive cancer risk exposure through independent oncogenic pathways, a scenario that has never been studied systematically.
The Role of IGF-1 LR3 in Combination Protocols
In the research and performance enhancement communities, IGF-1 LR3 is rarely used in isolation. It is frequently combined with other compounds in complex protocols that may include growth hormone, GH secretagogues, anabolic steroids, insulin, and other peptides. Each combination introduces unique interaction dynamics and risk profiles.
The most concerning combination is IGF-1 LR3 with exogenous insulin. Both compounds promote glucose uptake into cells, and their hypoglycemic effects are additive. Severe hypoglycemia from combined IGF-1/insulin use has resulted in documented fatalities in the bodybuilding community. This combination should be considered contraindicated outside of tightly controlled clinical settings with continuous glucose monitoring.
Combining LR3 with growth hormone creates a state of simultaneous GH excess and IGF-1 excess, mimicking some features of acromegaly. GH has direct effects that IGF-1 does not replicate (lipolysis, direct cartilage stimulation, diabetogenic effects through hepatic glucose output), so the combined exposure produces a broader spectrum of effects and side effects than either agent alone. The fluid retention, insulin resistance, and carpal tunnel syndrome associated with GH therapy may be amplified when combined with LR3.
Combining LR3 with tissue repair peptides like BPC-157 or TB-500 is a more common approach that raises fewer safety concerns, as these compounds operate through different receptor systems (BPC-157 through GH receptor upregulation and nitric oxide modulation; TB-500 through actin binding and cellular migration). The interactions are less well characterized but the pharmacological overlap is minimal, suggesting a lower risk of dangerous complementary effects.
Economic and Practical Comparison
Beyond pharmacology, practical considerations influence compound selection. IGF-1 LR3 is among the more expensive peptides on a per-dose basis, reflecting the complexity of recombinant protein production and purification. A typical 1 mg vial represents approximately 20-50 days of use at common doses (20-50 mcg/day). GH secretagogues like CJC-1295/Ipamorelin are generally less expensive per day of use and require less sophisticated storage (though they still require refrigeration after reconstitution). Oral compounds like MK-677 offer the greatest convenience (no injection required) at a competitive cost per day.
Storage requirements for LR3 are more demanding than for many peptides. The protein is sensitive to heat, light, agitation, and freeze-thaw cycles, all of which can cause irreversible denaturation and loss of activity. This means that practical potency may vary from theoretical potency depending on handling conditions during shipping, storage, and use, an uncontrollable variable that adds uncertainty to dosing precision.
Summary: Choosing Among Growth-Promoting Agents
Clinical Decision Framework
The choice among growth-promoting compounds depends on the specific goal, risk tolerance, and clinical context:
- Maximum anabolic potency (highest risk): IGF-1 LR3 or combined GH + IGF-1 protocols - reserved for research contexts with appropriate monitoring.
- Moderate anabolic support (moderate risk): GH secretagogues like CJC-1295/Ipamorelin or MK-677 - physiologically regulated, better safety data.
- Tissue repair focus: BPC-157 and TB-500 - different mechanism, complementary to anabolic compounds.
- Body composition (fat focus): AOD-9604, Fragment 176-191, or GLP-1 agents like semaglutide.
- Anti-aging/longevity: Epithalon, NAD+, Humanin, SS-31 - target aging mechanisms without the oncogenic concerns of IGF-1 elevation.
Use the dosing calculator for personalized guidance on specific compounds, and consult the drug comparison hub for additional head-to-head analyses.
Safety & Cancer Risk Considerations

What Are the Risks of IGF-1 LR3?
The safety profile of IGF-1 LR3 represents the central concern in any honest assessment of this compound. The same receptor activation that drives muscle growth - the PI3K/Akt/mTOR and MAPK/ERK cascades - also promotes cell proliferation, inhibits apoptosis, and enhances cell survival in virtually all cell types, including transformed (precancerous and cancerous) cells. This is not a theoretical concern. Large-scale epidemiological studies consistently associate higher circulating IGF-1 levels with increased cancer risk across multiple tumor types.
Because IGF-1 LR3 has never been subjected to formal clinical trials in humans for any indication, its safety profile must be extrapolated from three sources: clinical data with mecasermin (recombinant native IGF-1), epidemiological studies of endogenous IGF-1 levels and disease outcomes, and the clinical features of acromegaly (chronic endogenous IGF-1 excess).
Acute Safety Concerns
Hypoglycemia. This is the most common and most acutely dangerous adverse effect of IGF-1 therapy. In the primary mecasermin clinical trials of 71 subjects treated for a mean of 3.9 years, hypoglycemia was reported by 30 subjects (42%) at least once. Most episodes were mild to moderate, but 5 subjects experienced severe hypoglycemia requiring assistance from another person, and 4 subjects experienced hypoglycemic seizures or loss of consciousness.
IGF-1 LR3's extended half-life amplifies the hypoglycemia risk compared to mecasermin. While mecasermin's hypoglycemic effect typically peaks within a few hours of injection and can be managed by eating within 20 minutes of the dose, LR3's 20-30 hour duration means blood glucose lowering persists through the entire dosing interval, including during sleep when hypoglycemia is most dangerous. The risk is compounded by LR3's higher effective potency due to reduced IGFBP sequestration.
Injection site reactions. Pain, redness, and lipohypertrophy (localized fat accumulation) at injection sites are reported with mecasermin and expected with LR3. Chronic injection at the same site promotes lipohypertrophy, which can alter absorption kinetics unpredictably.
Headache and intracranial pressure. Increased intracranial pressure (pseudotumor cerebri/benign intracranial hypertension) has been reported with both GH and IGF-1 therapy. Symptoms include severe headache, visual changes, nausea, and vomiting. This is a serious adverse effect that requires prompt medical evaluation.
Fluid retention. Edema and water retention are common with IGF-1 therapy, reflecting the hormone's insulin-like effects on renal sodium handling. This can manifest as peripheral swelling, joint stiffness, and weight gain that does not represent true lean tissue accretion.
Chronic Safety Concerns: The Cancer Question
The relationship between IGF-1 and cancer is the most extensively studied and most concerning aspect of IGF-1 biology from a safety perspective. Multiple lines of evidence converge on the conclusion that sustained elevation of IGF-1 signaling increases cancer risk:
UK Biobank Cohort Study. The largest analysis to date, published in Cancer Research in 2020, examined circulating IGF-1 levels in over 300,000 UK Biobank participants with a median follow-up of 7 years. The study found positive associations between higher IGF-1 concentrations and risk of breast cancer (HR per SD increase = 1.10, 95% CI 1.07-1.14), prostate cancer (HR = 1.09, 95% CI 1.05-1.12), and colorectal cancer (HR = 1.07, 95% CI 1.02-1.11). These associations remained significant after adjustment for BMI, smoking, alcohol, physical activity, and other confounders (Murphy N, Knuppel A, Papadimitriou N, et al. Circulating insulin-like growth factor-1 and risk of total and 19 site-specific cancers: cohort study analyses from the UK Biobank. Cancer Res. 2020;80(20):4588-4598. PMID: 33067265).
EPIC-Heidelberg Study. Published in The Journal of Clinical Endocrinology & Metabolism in 2023, this case-cohort study within the European Prospective Investigation into Cancer and Nutrition found that higher IGF-1 was associated with increased risks of breast cancer (HR 1.25, 95% CI 1.06-1.47 per doubling) and prostate cancer (HR 1.31, 95% CI 1.09-1.57 per doubling). The prostate cancer association was particularly strong for advanced-stage disease (Tin Tin S, Reiter PL, et al. IGF-1 and risk of morbidity and mortality from cancer, cardiovascular diseases, and all causes in EPIC-Heidelberg. J Clin Endocrinol Metab. 2023;108(10):e1092-e1101. PMID: 37066827).
Mechanistic basis. The epidemiological associations are supported by a clear mechanistic rationale. The PI3K/Akt/mTOR pathway is one of the most commonly hyperactivated signaling cascades in human cancers. Activating mutations in PIK3CA (encoding the catalytic subunit of PI3K) are found in approximately 30% of breast cancers and many other tumor types. Loss of PTEN, the negative regulator of PI3K signaling, is also extremely common. IGF-1R activation feeds directly into this same pathway, and exogenous IGF-1 (especially long-acting variants like LR3) effectively provides a sustained exogenous "push" on a pathway that many incipient tumors are already predisposed to hyperactivate.
The IGF-1R also promotes cancer progression through anti-apoptotic signaling (Akt-mediated inactivation of pro-apoptotic factors), which protects cancer cells from immune surveillance and cytotoxic therapies. Indeed, IGF-1R has been actively pursued as an anti-cancer target, with several monoclonal antibodies and small molecule inhibitors developed against it for oncology indications.
Lessons from Acromegaly
Acromegaly provides a clinical "natural experiment" in chronic IGF-1 excess. Patients with uncontrolled acromegaly have persistently elevated IGF-1 levels (often 2-4 times the upper limit of normal) for years or decades before diagnosis. The consequences include:
- Cardiovascular disease: Hypertension affects 30-45% of acromegaly patients. Biventricular hypertrophy and diastolic dysfunction are present in up to 90% of cases at diagnosis. Without treatment, cardiovascular mortality is the leading cause of death.
- Metabolic disruption: Insulin resistance occurs in approximately 50% of patients, with overt diabetes mellitus in 10-30%. The paradox of IGF-1's insulin-like effects coexisting with insulin resistance reflects the complex metabolic consequences of chronic GH/IGF-1 excess.
- Organomegaly: Enlargement of the liver, kidneys, spleen, and other visceral organs is characteristic. Cardiac enlargement contributes to the cardiomyopathy. Soft tissue hypertrophy causes macroglossia (tongue enlargement), carpal tunnel syndrome, and skin thickening.
- Cancer risk: Colonoscopy findings in acromegaly patients show increased colon length, mucosal hypertrophy, and elevated polyp prevalence. Up to 25% of polyps are right-sided (a location associated with higher malignant potential) and recur within 3 years. Overall cancer mortality is increased, particularly for colorectal cancer.
- Reduced life expectancy: Untreated acromegaly reduces life span by approximately 10 years (roughly 30% of expected remaining life), primarily through cardiovascular, cerebrovascular, and respiratory complications.
While the degree of IGF-1 elevation in acromegaly is typically greater and more sustained than what exogenous LR3 administration would produce (at commonly discussed doses), the acromegaly data establishes a clear biological principle: chronic IGF-1 excess has serious, multi-system health consequences. The question for LR3 users is not whether these risks exist, but at what level and duration of exposure they become clinically significant. That question remains unanswered by current evidence.
The IGF-1R as a Cancer Target: Implications for IGF-1 Administration
Perhaps the most telling indicator of the relationship between IGF-1 signaling and cancer is the pharmaceutical industry's extensive investment in developing IGF-1R inhibitors as anti-cancer therapies. Multiple monoclonal antibodies targeting IGF-1R (figitumumab, dalotuzumab, ganitumab, cixutumumab, robatumumab) and small molecule IGF-1R tyrosine kinase inhibitors (linsitinib, BMS-754807) have been developed and tested in clinical trials across numerous tumor types including non-small cell lung cancer, breast cancer, pancreatic cancer, and sarcoma.
While these agents have generally produced disappointing results in unselected patient populations (leading to the termination of most clinical development programs), the scientific rationale for targeting IGF-1R in cancer remains sound. The pathway is frequently activated in cancer, and preclinical models consistently show that IGF-1R inhibition reduces tumor growth, enhances sensitivity to chemotherapy, and impairs metastasis. The clinical failures have been attributed to lack of patient selection biomarkers, compensatory signaling through hybrid receptors and the insulin receptor, and inadequate pathway suppression rather than to an invalid target hypothesis.
The relevance for IGF-1 LR3 users is straightforward: the same receptor that pharmaceutical companies have spent billions of dollars trying to block in cancer patients is the receptor that LR3 is designed to activate. While this does not mean that LR3 use will cause cancer, it means that LR3 use creates a biochemical environment that, based on extensive preclinical and clinical research, is favorable for tumor initiation and progression. For individuals with existing precancerous lesions (which many adults have without knowing it, particularly colon polyps and prostatic intraepithelial neoplasia), sustained IGF-1R activation could potentially accelerate the transition from premalignant to malignant disease.
IGF-1 and the Longevity Paradox
An additional consideration comes from the longevity literature, which presents what appears to be a paradox: while IGF-1 promotes growth, tissue maintenance, and muscle preservation (all seemingly beneficial for healthspan), reduced IGF-1 signaling is consistently associated with extended lifespan across species from yeast to mice. Dwarf mice with reduced GH/IGF-1 signaling live 30-65% longer than their normal counterparts. Centenarian humans show functional mutations in the IGF-1R and downstream signaling components that partially reduce pathway activity.
This creates a genuine biological tension. IGF-1 is pro-growth and pro-survival at the cellular level, which promotes tissue maintenance and repair in the short term. But at the organismal level, the cumulative effect of enhanced cellular growth and survival over decades appears to increase cancer risk and accelerate certain aspects of aging (including the accumulation of senescent cells that resist apoptosis, potentially in an IGF-1-dependent manner). Compounds like FOXO4-DRI aim to selectively eliminate senescent cells, addressing one consequence of the growth-promoting signaling that IGF-1 drives.
For individuals weighing the use of IGF-1 LR3, this longevity data provides important context. The muscle growth and tissue repair benefits of enhanced IGF-1 signaling may come at the cost of accelerated cellular aging and increased long-term disease risk. Whether this tradeoff is acceptable depends on individual priorities, risk tolerance, and the specific clinical context, but it should be made with full awareness of the evidence on both sides.
Neurological and Cognitive Effects
IGF-1's effects on the central nervous system deserve mention in a safety context because they represent both a potential benefit and a potential risk. IGF-1 crosses the blood-brain barrier and supports neuronal survival, synaptogenesis, and myelination. Low IGF-1 levels are associated with cognitive decline in aging, and several studies have explored IGF-1 as a potential neuroprotective therapy.
The 2024 study by researchers examining intranasal LR3-IGF-1 in a 5XFAD mouse model of Alzheimer's disease found that the treatment promoted amyloid plaque remodeling in the cerebral cortex, with in vitro experiments showing enhanced microglial uptake of amyloid-beta peptide. However, the same study found that the treatment failed to preserve cognitive function, and treated mice showed some behavioral changes (PMID: 39610283). This mixed result illustrates the complexity of IGF-1's CNS effects: it can influence pathology without necessarily improving function, and the consequences of sustained IGF-1R activation in brain tissue remain incompletely understood.
Cognitive peptides like Semax, Selank, and Dihexa offer alternative approaches to cognitive support through mechanisms that don't carry the same oncogenic concerns as IGF-1R activation. Pinealon and P21 also act on neurological pathways through distinct receptor systems.
Dose-Dependent and Duration-Dependent Risk Stratification
When evaluating cancer risk from IGF-1 elevation, the concepts of dose-dependence and duration-dependence are critical. The epidemiological data cited above (UK Biobank, EPIC-Heidelberg) examine endogenous IGF-1 variation within the normal physiological range. The hazard ratios of 1.07-1.31 per standard deviation or doubling of IGF-1 concentration reflect the risk associated with being at the upper vs. lower end of the normal distribution. Exogenous LR3 administration could potentially push total IGF-1R activation well above the normal range, into territory where the risk amplification is unknown but likely greater than these observational estimates suggest.
Duration of exposure is equally important. The acromegaly literature shows that the health consequences of IGF-1 excess develop over years, not weeks. Cardiovascular remodeling, metabolic disruption, and clinically apparent tumors typically emerge after years of uncontrolled disease. This might suggest that short-cycle LR3 use (4-6 weeks) poses lower risk than chronic exposure. However, the cancer biology literature also shows that even brief exposures to carcinogenic stimuli can initiate the multistep process of carcinogenesis by causing genetic mutations, epigenetic changes, or selection pressure favoring pre-existing mutant clones. Once initiated, a cancer may progress even after the initiating stimulus is removed.
The practical implication is that while shorter cycles at lower doses are undoubtedly safer than longer cycles at higher doses, there is no known duration or dose that can be declared "safe" from a cancer risk perspective. This is a fundamental limitation of any compound that acts as a mitogen (cell proliferation promoter), and it applies to IGF-1 LR3 as surely as it applies to any growth factor.
Cardiovascular Safety Considerations
Beyond cancer, the cardiovascular implications of IGF-1 LR3 deserve careful attention. IGF-1 has complex effects on the cardiovascular system that depend on dose, duration, and the baseline cardiac state:
Cardiac hypertrophy. IGF-1 directly stimulates cardiomyocyte hypertrophy through the same PI3K/Akt pathway that drives skeletal muscle growth. In physiological amounts and in the context of exercise training, this produces beneficial "athlete's heart" remodeling: symmetric hypertrophy with preserved or enhanced diastolic function. In pathological excess (as in acromegaly), the same pathway drives concentric hypertrophy with diastolic dysfunction, fibrosis, and ultimately cardiomyopathy. The threshold between physiological and pathological cardiac remodeling in the context of exogenous IGF-1 administration is unknown.
Fluid retention and blood pressure. IGF-1 promotes renal sodium and water retention, which increases blood volume and can elevate blood pressure. In the mecasermin trials, edema and increased blood pressure were reported adverse events. For individuals with borderline or established hypertension, LR3's fluid-retaining effects could push blood pressure into dangerous territory, particularly in the setting of exercise-induced cardiac demand.
Electrolyte disturbances. The fluid retention and glucose-lowering effects of IGF-1 can secondarily affect electrolyte balance, particularly potassium homeostasis. Insulin-like glucose uptake is accompanied by intracellular potassium shifting, which can cause transient hypokalemia. In the setting of concurrent exercise (with potassium loss through sweat), this could theoretically contribute to cardiac arrhythmia risk, though this specific scenario has not been studied.
Vascular effects. IGF-1 promotes vascular smooth muscle proliferation and has both pro-angiogenic and pro-atherogenic properties depending on the context. In the short term, IGF-1 may promote beneficial endothelial function through nitric oxide synthesis. In the long term, it may contribute to intimal hyperplasia and accelerated atherosclerosis, effects well-documented in acromegaly patients.
Metabolic Safety: Beyond Hypoglycemia
While acute hypoglycemia receives the most attention, the metabolic effects of IGF-1 LR3 extend beyond glucose lowering:
Insulin resistance paradox. Acutely, IGF-1 has insulin-like glucose-lowering effects. But chronically, sustained IGF-1R activation can paradoxically promote insulin resistance. This occurs through several mechanisms: IGF-1R and IR share downstream signaling components, and chronic activation can desensitize shared pathway elements (particularly IRS-1 phosphorylation); IGF-1 promotes growth of insulin-resistant tissues (visceral organ enlargement); and the compensatory hyperinsulinemia triggered by IGF-1-induced hypoglycemia may itself promote insulin resistance over time. In acromegaly, approximately 50% of patients develop insulin resistance and 10-30% develop overt diabetes mellitus, despite having both elevated GH (which is diabetogenic) and elevated IGF-1 (which is insulin-sensitizing in acute settings).
Lipid metabolism. IGF-1's effects on lipid metabolism are complex and context-dependent. In GH-deficient patients, normalizing IGF-1 through GH replacement improves lipid profiles (reducing LDL cholesterol and triglycerides). However, exogenous IGF-1 without concurrent GH has different metabolic effects because GH has direct lipolytic properties that IGF-1 lacks. LR3 administration without GH may promote insulin-like lipogenic effects, potentially increasing adiposity in certain compartments even while promoting lean mass growth.
Thyroid function. The GH-IGF-1 axis interacts with thyroid hormone metabolism. GH increases the conversion of T4 to T3 (the active thyroid hormone) by stimulating hepatic 5'-deiodinase. Exogenous IGF-1 that suppresses endogenous GH secretion through feedback inhibition may reduce this conversion, potentially causing a relative T3 deficiency. Clinical significance in the context of short-term LR3 cycles is uncertain, but monitoring thyroid function is reasonable during extended use.
Suppression of Endogenous IGF System
An underappreciated risk of exogenous IGF-1 LR3 administration is suppression of the endogenous GH-IGF-1 axis. As demonstrated in the Lok et al. guinea pig infusion study, systemic LR3 administration reduced circulating levels of endogenous IGF-1, IGF-2, and multiple IGFBPs. This occurs through negative feedback: exogenous IGF-1 receptor activation at the hypothalamic and pituitary level suppresses GH secretion, which in turn reduces hepatic IGF-1 production.
This means that cessation of LR3 use is followed by a period of relative IGF-1 deficiency as the suppressed axis recovers. The duration and severity of this suppression depends on the dose and duration of LR3 use, but it parallels the well-described hypothalamic-pituitary suppression seen with exogenous steroid use. For individuals using LR3 for muscle maintenance, this rebound deficiency could potentially accelerate muscle loss during the recovery period, creating a problematic dependency cycle.
Product Quality and Contamination Risk
Because IGF-1 LR3 is not manufactured under pharmaceutical GMP (Good Manufacturing Practice) standards for human therapeutic use, products available through research chemical suppliers and compounding pharmacies vary significantly in purity, potency, and sterility. Recombinant protein production involves complex expression systems (typically E. coli or mammalian cell culture), and inadequate purification can leave endotoxin contamination, misfolded protein variants, or truncation products in the final material.
The consequences of injecting contaminated or impure protein products include injection site infections, immune reactions (antibody formation that can neutralize both the exogenous product and endogenous IGF-1), and unpredictable potency leading to either underdosing or dangerous overdosing. This practical safety concern is separate from the pharmacological risks of IGF-1 itself but adds another layer of hazard for anyone using non-pharmaceutical-grade material.
Safety Summary
IGF-1 LR3 carries significant acute and chronic safety risks that are inherent to its mechanism of action. Acute risks include hypoglycemia (potentially severe), fluid retention, headache, and injection site reactions. Chronic risks include increased cancer susceptibility (supported by large epidemiological studies), potential organ enlargement, metabolic disruption, and suppression of the endogenous IGF system. The absence of controlled human clinical trial data for LR3 specifically means that the therapeutic index (the margin between effective and harmful doses) is unknown. Medical supervision, including regular monitoring of blood glucose, IGF-1 levels, and age-appropriate cancer screening, is essential for anyone using this compound. The free assessment at FormBlends can help determine whether peptide therapy is appropriate for your specific situation.
Dosing Considerations

What Are the Dosing Considerations for IGF-1 LR3?
IGF-1 LR3 dosing protocols are derived entirely from preclinical data, extrapolation from mecasermin clinical experience, and anecdotal reports from the research and bodybuilding communities. No dose-finding studies, phase I safety trials, or randomized efficacy trials have been conducted with LR3 in humans. Every dosing recommendation that follows carries this essential caveat: the therapeutic index is unknown, and individual responses can vary substantially.
With that important disclaimer established, here is what the available evidence and clinical extrapolation suggest about responsible dosing approaches.
Dose Ranges Reported in the Literature and Community
The most commonly referenced dose ranges for IGF-1 LR3 in the research community fall into three tiers:
| Tier | Daily Dose | Context | Risk Level |
|---|---|---|---|
| Conservative / Beginner | 20-30 mcg/day | Initial exposure assessment | Lower (but not negligible) |
| Moderate | 40-60 mcg/day | Commonly reported effective range | Moderate |
| High / Advanced | 80-100 mcg/day | Upper range reported anecdotally | High (exceeds evidence base) |
Doses above 50-60 mcg/day lack meaningful research support and are associated with substantially increased risk of adverse effects, particularly hypoglycemia. The mcg-level dosing reflects LR3's high potency; by comparison, mecasermin is dosed at 0.04-0.12 mg/kg twice daily (equivalent to roughly 2,000-8,000 mcg/day for an 80 kg adult), but mecasermin is subject to extensive IGFBP sequestration that dramatically reduces its effective free concentration.
Administration Route and Timing
Subcutaneous injection is the most common administration route for IGF-1 LR3. Injection is typically performed in the abdominal subcutaneous tissue using insulin syringes (29-31 gauge, 0.5-1.0 mL). Rotation of injection sites is important to prevent lipohypertrophy.
Intramuscular injection is sometimes used with the rationale that it might increase local muscle IGF-1 receptor exposure. However, given LR3's long half-life and systemic distribution, any site-specific advantage from intramuscular injection is likely minimal. IGF-1 DES is better suited for site-specific intramuscular approaches due to its short half-life and rapid local action.
Timing relative to meals. This is critically important for safety. Because IGF-1 LR3 can cause significant hypoglycemia, injection should be performed shortly before or immediately after consuming a meal containing both carbohydrates and protein. Post-workout timing (within 30-60 minutes of completing resistance training) is commonly recommended in the research community, as this coincides with a period of elevated IGF-1 receptor sensitivity in exercised muscles and the natural post-exercise feeding window.
The carbohydrate component of the post-injection meal serves a dual purpose: providing substrate for the insulin-like glucose uptake effect of IGF-1, and stimulating endogenous insulin release that, together with IGF-1's effects, supports anabolic signaling in muscle. A meal providing 30-50 grams of carbohydrate and 20-40 grams of protein within 30 minutes of injection is a commonly cited guideline.
Cycle Structure
Continuous, indefinite use of IGF-1 LR3 is universally discouraged in the research literature due to concerns about receptor desensitization, endogenous axis suppression, and cumulative cancer risk. Cycling protocols typically follow a 4-6 week "on" period followed by an equal or longer "off" period:
- Conservative cycle: 4 weeks on, 4-6 weeks off. This allows some recovery of the endogenous GH-IGF-1 axis and limits cumulative exposure.
- Standard cycle: 4-6 weeks on, 4-8 weeks off. The most commonly reported protocol in community literature.
- Extended cycle: 6-8 weeks on. Reported anecdotally but associated with greater axis suppression and higher cumulative risk. Not recommended without close medical monitoring.
During the off-cycle period, some practitioners transition to CJC-1295/Ipamorelin or other GH secretagogues to support endogenous axis recovery while maintaining some degree of GH-IGF-1 axis stimulation through physiological mechanisms. The dosing calculator can help with planning transition protocols between compounds.
Blood Glucose Management Protocol
Given that hypoglycemia is the most common and most acutely dangerous adverse effect of IGF-1 therapy, a detailed blood glucose management protocol is essential for anyone using IGF-1 LR3. The following framework draws on the mecasermin prescribing information, diabetes management guidelines, and clinical experience with insulin-like hypoglycemic agents.
Pre-injection assessment. Blood glucose should be checked before each injection. If blood glucose is below 70 mg/dL (3.9 mmol/L), the injection should be postponed until glucose has been raised with food. Injecting LR3 in the setting of existing hypoglycemia can produce dangerous blood sugar drops.
Meal requirements. Every injection must be accompanied by a meal. The mecasermin prescribing information recommends injection within 20 minutes of a meal. For LR3, the same principle applies, with the post-injection meal ideally consumed within 15-30 minutes. The meal should contain at minimum 30 grams of complex carbohydrates (to provide sustained glucose availability during LR3's prolonged action) and 20-30 grams of protein (to support the anabolic signaling that the injection is intended to promote).
Post-injection monitoring. During the first week of use (or with any dose increase), blood glucose should be checked at 1 hour, 2 hours, and 4 hours post-injection to characterize the individual's glucose response curve. A continuous glucose monitor (CGM) such as FreeStyle Libre, Dexcom G7, or similar device provides far more comprehensive data and is strongly recommended. CGM data can reveal nocturnal hypoglycemia that finger-stick monitoring would miss entirely.
Nocturnal hypoglycemia prevention. Because LR3's half-life extends through the night following a daytime injection, nocturnal hypoglycemia is a real risk. A bedtime snack containing slow-release carbohydrates (e.g., oatmeal, whole grain bread with peanut butter) helps maintain blood glucose overnight. CGM with low-glucose alerts is the gold standard for nocturnal safety monitoring.
Emergency hypoglycemia management. Users should always have rapid-acting glucose sources immediately available: glucose tablets (15 grams glucose per dose), fruit juice (120 mL/4 oz provides approximately 15 grams glucose), or glucose gel. The "Rule of 15" applies: consume 15 grams of fast-acting carbohydrate, wait 15 minutes, re-check glucose, and repeat if glucose remains below 70 mg/dL. For severe hypoglycemia (confusion, inability to eat, loss of consciousness), someone trained in glucagon administration should be present. Injectable glucagon kits or intranasal glucagon (Baqsimi) should be available in the household of anyone using IGF-1 LR3.
Injection Technique and Site Selection
Proper injection technique is important for both safety and consistent dosing. Subcutaneous injection delivers LR3 into the fatty layer beneath the skin, where it is absorbed into the systemic circulation at a rate determined by local blood flow and subcutaneous tissue characteristics.
Site selection. Common subcutaneous injection sites include the abdominal wall (avoiding a 2-inch radius around the navel), the anterior thigh, and the back of the upper arm. Abdominal injections generally produce the most consistent absorption. Each injection should use a different site within the chosen area, rotating in a systematic pattern to prevent lipohypertrophy (localized fat accumulation) or lipoatrophy (localized fat loss) that can develop with repeated injection at the same spot.
Technique. Using an insulin syringe (29-31 gauge, 6 mm or 8 mm needle length), pinch the skin at the chosen site, insert the needle at a 45-90 degree angle (depending on subcutaneous tissue thickness), inject slowly, hold for 5-10 seconds after injection to prevent leakback, then release the skin pinch and withdraw. Aspiration (pulling back on the plunger to check for blood return) is not recommended for subcutaneous injection as it provides no safety benefit and increases discomfort.
Intramuscular technique. For those choosing intramuscular administration (typically targeting specific muscle groups post-workout), a longer needle may be needed depending on subcutaneous fat thickness. Common IM injection sites include the vastus lateralis (outer quadriceps), deltoid, and gluteus medius. IM injection may produce slightly faster absorption than subcutaneous but with more local discomfort and greater variability in absorption rate.
Reconstitution and Storage
IGF-1 LR3 is typically supplied as a lyophilized (freeze-dried) powder, usually in 1 mg (1,000 mcg) vials. Reconstitution requires careful technique to preserve the protein's biological activity:
Reconstitution solvent: Sterile bacteriostatic water (containing 0.9% benzyl alcohol as a preservative) is the recommended solvent. Normal saline (0.9% NaCl) or sterile water for injection can also be used, but without the bacteriostatic preservative, the reconstituted solution must be used within a shorter timeframe (24-48 hours if refrigerated).
Reconstitution technique: Add the solvent slowly, directing the stream down the inside wall of the vial rather than directly onto the lyophilized powder. Allow the powder to dissolve gradually; gentle swirling is acceptable, but vigorous shaking should be avoided as it can denature the protein and reduce potency. A 1 mg vial reconstituted with 1 mL of bacteriostatic water yields a concentration of 1,000 mcg/mL; reconstitution with 2 mL yields 500 mcg/mL. The choice depends on the desired injection volume precision.
Storage: Lyophilized (unreconstituted) LR3 should be stored at -20C (freezer) for long-term stability or 2-8C (refrigerator) for shorter-term storage. Reconstituted solution should be stored at 2-8C and used within 4 weeks (with bacteriostatic water) or within 24-48 hours (without preservative). Avoid freezing reconstituted solution, as freeze-thaw cycles can denature the protein. Protect from light.
Monitoring Requirements
Anyone using IGF-1 LR3 should have medical supervision and regular monitoring. At minimum, the following parameters should be assessed:
- Blood glucose: Frequent self-monitoring during the initial dosing period, then at least daily during the cycle. A continuous glucose monitor (CGM) provides the most comprehensive safety data, particularly for detecting nocturnal hypoglycemia.
- Serum IGF-1 levels: Baseline and during treatment to assess the degree of IGF-1 elevation and calibrate dosing.
- HbA1c: At baseline and after each cycle to monitor cumulative glycemic impact.
- Complete metabolic panel: Liver and kidney function tests, as these organs are primary sites of IGF-1 clearance and may be affected by sustained receptor activation.
- Age-appropriate cancer screening: Given the epidemiological associations between IGF-1 and cancer, current cancer screening guidelines should be followed rigorously. PSA testing for men over 50 (or earlier with risk factors) and mammographic screening for women per applicable guidelines.
- Cardiac assessment: Baseline echocardiography is prudent, particularly for individuals with cardiovascular risk factors, given the association between chronic IGF-1 excess and cardiomyopathy.
Contraindications
IGF-1 LR3 should not be used in the following circumstances:
- Active malignancy or history of cancer (particularly breast, prostate, or colorectal)
- Active or suspected neoplasia of any type
- Conditions predisposing to hypoglycemia (insulinoma, adrenal insufficiency, severe liver disease)
- Concurrent use of insulin or sulfonylurea medications (greatly increased hypoglycemia risk)
- Pregnancy or breastfeeding (growth factor effects on fetal/neonatal development are unpredictable)
- Uncontrolled diabetes mellitus
- Known hypersensitivity to IGF-1 or any excipients in the formulation
- Closed epiphyses in children/adolescents receiving IGF-1 for growth promotion (reduced efficacy)
- Severe cardiac disease or heart failure (risk of fluid retention and cardiac complications)
Clinical Guidance
The single most important safety measure for IGF-1 LR3 users is consistent meal timing around injections. Never inject on an empty stomach, never skip the post-injection meal, and always have a rapid-acting carbohydrate source available (glucose tablets, juice) to treat hypoglycemic episodes. Nocturnal injection should be avoided due to the risk of hypoglycemia during sleep. These precautions, drawn directly from the mecasermin prescribing information and clinical experience, apply with even greater urgency to LR3 given its enhanced potency and duration. For personalized assessment of whether IGF-1 LR3 is appropriate for your goals, the free assessment is a recommended starting point.
Interactions with Other Compounds
IGF-1 LR3's pharmacological profile creates several important interaction considerations:
- Insulin: Combined use of IGF-1 LR3 and insulin dramatically increases hypoglycemia risk and is considered extremely dangerous. The insulin-like effects of LR3 are additive with exogenous insulin, potentially causing severe, prolonged hypoglycemia that can result in seizures, coma, or death.
- Growth hormone: Combined GH + LR3 use can produce supraphysiological activation of both direct GH signaling and IGF-1 receptor signaling simultaneously, amplifying both anabolic effects and side effects (particularly fluid retention, insulin resistance, and cancer risk).
- GH secretagogues: CJC-1295/Ipamorelin, MK-677, and similar compounds increase endogenous GH and IGF-1. Combined use with exogenous LR3 creates additive IGF-1 receptor stimulation. If used together, lower doses of each component are recommended.
- Oral hypoglycemic agents: Metformin, sulfonylureas, and other glucose-lowering medications may interact with LR3's hypoglycemic effects. Dose adjustment of diabetes medications may be necessary.
- GLP-1 receptor agonists: Compounds like semaglutide and tirzepatide affect glucose metabolism and appetite. Combined use with IGF-1 LR3 could create complex glucose dynamics requiring careful monitoring.
Understanding Dose-Response Relationships
One of the fundamental challenges with IGF-1 LR3 dosing is the absence of formal dose-response data in humans. In pharmacology, establishing a dose-response curve requires testing multiple doses in a controlled setting to determine the minimum effective dose, the dose producing maximum benefit, and the dose at which adverse effects become unacceptable. None of this has been done for LR3.
What we can infer from the broader IGF-1 literature suggests a relatively narrow therapeutic window. The mecasermin experience shows that even at carefully calibrated doses with meal timing, 42% of patients experience hypoglycemia. The epidemiological data shows that cancer risk increases linearly with IGF-1 concentration, without a clear threshold below which risk is zero. These observations suggest that the margin between an "effective" and a "harmful" dose of IGF-1R activation may be quite small.
In preclinical models, the dose-response relationship for IGF-1's anabolic effects on muscle follows a saturable curve. At low doses, increasing IGF-1 produces proportional increases in protein synthesis and satellite cell activation. At moderate doses, the response begins to plateau as downstream signaling effectors (particularly mTOR and its targets) approach maximum activation. At high doses, additional IGF-1 produces minimal further anabolic benefit but continues to increase adverse effects (hypoglycemia, fluid retention, and mitogenic stimulation of non-muscle tissues).
For LR3 specifically, the dose range that produces maximal muscle anabolism relative to systemic side effects is unknown. The commonly cited 20-60 mcg/day range is based on anecdotal reports and back-of-envelope calculations comparing LR3's effective potency to mecasermin's established doses, not on systematic dose-finding. Individual variation in IGF-1R expression density, IGFBP milieu (even LR3 interacts with IGFBPs to some degree at high concentrations), body composition, insulin sensitivity, and genetic polymorphisms in the IGF-1 signaling pathway all contribute to unpredictable individual responses.
Sex-Based Dosing Differences
Men and women differ in several aspects of IGF-1 biology that should influence dosing considerations. Women generally have lower circulating IGF-1 levels than men of the same age (by approximately 10-15%), different IGFBP profiles (higher IGFBP-1, influenced by estrogen status), and may have different tissue-specific IGF-1R expression patterns. The dose-response to exogenous IGF-1 has not been systematically compared between sexes.
Community reports suggest that women typically use lower doses (10-20 mcg/day) than men (20-60 mcg/day), though the basis for this difference is more cautionary principle than evidence-based dosing. Given the absence of sex-specific pharmacokinetic data for LR3, the conservative approach of starting at the lower end of any dose range and titrating based on response and side effects seems particularly important for female users.
Estrogen status may also influence the safety profile. Premenopausal women have higher IGFBP-1 levels (stimulated by estrogen), which could theoretically provide some degree of buffering even for LR3 (which still retains some residual IGFBP interaction at high circulating concentrations). Postmenopausal women or women on hormonal contraceptives may have altered IGFBP profiles that affect LR3's effective bioavailability differently.
Special Populations and Age Considerations
The risk-benefit calculation for IGF-1 LR3 varies substantially by age and health status:
Young adults (18-30): This population already has near-peak endogenous IGF-1 levels, high satellite cell numbers, and strong mTOR signaling in response to exercise. The marginal benefit of exogenous LR3 is likely smallest in this group, while the exposure to prolonged IGF-1R activation begins accumulating at the youngest age, creating the largest lifetime cancer risk burden. From a risk-benefit perspective, this group has the least to gain and potentially the most to lose.
Middle-aged adults (40-55): Endogenous IGF-1 has declined 20-30% from peak, satellite cell function is beginning to decline, and the anabolic response to exercise is somewhat attenuated. The potential benefit of IGF-1 supplementation increases relative to younger adults, but cancer risk also increases with age (independently of IGF-1), creating a situation where even modest IGF-1R activation contributes to a higher baseline cancer risk.
Older adults (60+): This population has the most to gain from IGF-1 signaling restoration in terms of combating sarcopenia, frailty, and age-related functional decline. However, they also have the highest baseline cancer risk, the highest prevalence of occult premalignant lesions, and the greatest sensitivity to the metabolic effects (hypoglycemia, fluid retention) that IGF-1 excess produces. The risk-benefit balance is most nuanced in this group and most dependent on individual health status and cancer screening results.
For all age groups, the alternative of GH secretagogue therapy offers a way to increase IGF-1 within the physiological range and with preserved regulatory feedback, potentially capturing some of the benefits of IGF-1 restoration without the risks of unregulated receptor activation that LR3 creates.
Post-Cycle Recovery and Axis Restoration
After discontinuing IGF-1 LR3, the suppressed GH-IGF-1 axis requires time to recover. During LR3 use, the exogenous IGF-1R activation feeds back to suppress hypothalamic GHRH release and pituitary GH secretion. This reduces endogenous hepatic IGF-1 production. When LR3 is discontinued, there is a transient period during which both exogenous (cleared) and endogenous (suppressed) IGF-1 are low, creating a relative IGF-1 deficiency that may produce symptoms including fatigue, reduced exercise recovery, mood changes, and accelerated muscle protein breakdown.
The duration of axis suppression depends on the dose and duration of LR3 use. Based on analogies with other exogenous hormone axes (testosterone, corticosteroids), a 4-6 week LR3 cycle is estimated to produce axis suppression lasting 2-4 weeks after discontinuation. Longer cycles or higher doses may produce more prolonged suppression.
Several strategies have been proposed to facilitate axis recovery:
- GH secretagogue transition: Starting a GH secretagogue like sermorelin or CJC-1295/Ipamorelin during the off-cycle period can stimulate endogenous GH pulsatility and support IGF-1 recovery while maintaining some degree of growth hormone axis activation.
- Adequate sleep optimization: Since GH secretion is predominantly nocturnal (during deep sleep), optimizing sleep quality and duration supports natural axis recovery. DSIP (Delta Sleep Inducing Peptide) has been investigated for its sleep-promoting properties, though evidence for its use in this specific context is limited.
- Resistance training continuation: Exercise-induced local IGF-1 (MGF) production occurs independently of the systemic GH-IGF-1 axis. Continuing a structured resistance training program during the off-cycle maintains local anabolic signaling even while the systemic axis recovers.
- Nutritional support: Adequate protein intake (1.6-2.2 g/kg/day), sufficient caloric intake (avoiding deficits that suppress GH), and micronutrient adequacy (particularly zinc, magnesium, and vitamin D, which influence GH and IGF-1 production) all support natural axis function.
Quality Assurance Considerations
Because IGF-1 LR3 is not manufactured under pharmaceutical GMP standards for human use, product quality varies enormously across suppliers. Key quality parameters that affect safety and efficacy include:
Protein purity. High-quality LR3 should be at least 95% pure (ideally >98%) as determined by reversed-phase HPLC. Lower purity products may contain truncated protein fragments, misfolded variants, or host cell proteins from the expression system that can cause immune reactions or unpredictable biological effects.
Endotoxin content. Bacterial endotoxins (lipopolysaccharides) are potent pyrogens and immune activators. Products expressed in E. coli must undergo rigorous endotoxin removal. Endotoxin levels should be below 1 EU/mg (endotoxin units per milligram) for injectable products. Products with high endotoxin content can cause fever, injection site inflammation, and systemic inflammatory responses.
Correct folding. IGF-1 LR3 contains three disulfide bonds that must be correctly formed for biological activity. Incorrect disulfide pairing produces misfolded variants that may have reduced potency, altered receptor specificity, or increased immunogenicity. Bioactivity testing (typically using cell proliferation assays) should confirm that the product has the expected biological potency.
Sterility. Products intended for injection must be sterile. Lyophilized products produced under non-GMP conditions should be handled with strict aseptic technique during reconstitution, and users should be aware that non-pharmaceutical products may not have undergone terminal sterilization or sterility testing.
Identity confirmation. Mass spectrometry (MALDI-TOF or ESI-MS) should confirm the correct molecular weight (approximately 9,111 Da for LR3), and amino acid analysis or peptide mapping should verify the primary sequence. Products that don't provide analytical data (certificates of analysis) should be viewed with skepticism.
Peptide content verification. Some products marketed as IGF-1 LR3 may contain different quantities than labeled, or may contain entirely different peptides. Independent third-party testing through services that offer peptide identification and quantitation provides additional assurance beyond the supplier's own quality documentation.
Excipient safety. Lyophilized peptide products may contain excipients such as mannitol, trehalose, or sucrose as cryoprotectants (which protect the protein during freeze-drying), and acetic acid or sodium acetate as pH buffers. These are generally safe at the small quantities present in a reconstituted injection. However, some non-pharmaceutical products may contain unusual or undisclosed excipients that could cause allergic reactions or injection site irritation. Users with known sensitivities should verify the full composition of any product before use.
The practical implication is that individuals using IGF-1 LR3 should source products from reputable suppliers who provide comprehensive certificates of analysis including purity (HPLC), identity (mass spectrometry), endotoxin testing, and bioactivity data. The cost of analytical testing is reflected in price; products that are dramatically cheaper than competitors may have cut corners on quality control.
The Legal and Regulatory Environment
The regulatory status of IGF-1 LR3 varies by jurisdiction and intended use, creating a complex legal environment for researchers, clinicians, and individuals:
United States. IGF-1 LR3 is not FDA-approved for any therapeutic indication. It is classified as a research chemical and can be legally purchased for research purposes from chemical suppliers. However, it is a controlled substance under the Anti-Drug Abuse Act's provisions regarding human growth hormone (HGH) analogs. The Anabolic Steroid Control Act of 2004 added certain "prohormones" to the controlled substance schedule, and while IGF-1 LR3 is not specifically listed as an anabolic steroid, its legal status for human use outside of a legitimate research or clinical context is ambiguous. Distribution for human consumption without FDA approval would be illegal.
Compounding pharmacies in some states may prepare IGF-1 LR3 under physician prescription as part of a compounded medication protocol. The legality and regulatory oversight of such compounding varies by state and is subject to both state pharmacy board regulations and federal FDA enforcement priorities.
World Anti-Doping Agency (WADA). IGF-1 and all of its analogs, including LR3, are prohibited substances under WADA's Prohibited List (Category S2: Peptide Hormones, Growth Factors, Related Substances and Mimetics). Athletes subject to WADA testing who use IGF-1 LR3 face suspension from competition. Detection methods for IGF-1 analogs have improved significantly in recent years, using mass spectrometry-based approaches that can distinguish exogenous from endogenous IGF-1.
International variation. In some jurisdictions, IGF-1 LR3 may be available as a prescription medication through compounding pharmacies, while in others it may be entirely unregulated or completely prohibited. Users should understand the specific legal framework in their jurisdiction before obtaining or using this compound.
Emerging Research Directions
Several lines of ongoing research may reshape our understanding of IGF-1 LR3's potential and limitations:
Targeted delivery systems. Nanoparticle-based delivery, tissue-homing peptide conjugates, and hydrogel depots are being investigated to deliver IGF-1 locally to muscle tissue without systemic exposure. If successful, these approaches could capture the anabolic benefits of IGF-1R activation in muscle while minimizing the cancer risk, metabolic disruption, and organ enlargement associated with systemic exposure. Preclinical work with injectable hydrogels loaded with IGF-1 has shown sustained local release with enhanced muscle regeneration in injury models.
Selective IGF-1R modulators. Analogous to selective estrogen receptor modulators (SERMs) that produce tissue-specific effects, researchers are investigating whether IGF-1 analogs can be engineered to produce biased signaling - activating the PI3K/Akt pathway (anabolic) preferentially over the MAPK/ERK pathway (mitogenic) or vice versa. If the growth-promoting and cancer-promoting activities of IGF-1R activation can be pharmacologically separated, it would fundamentally change the risk-benefit equation for IGF-1-based therapies.
Gene therapy approaches. Building on the transgenic mouse work demonstrating that muscle-specific IGF-1 overexpression preserves muscle mass during aging without systemic effects, researchers are exploring AAV (adeno-associated virus)-mediated IGF-1 gene delivery to skeletal muscle. This approach could provide sustained, localized IGF-1 production at physiological levels, replicating the protective effects seen in transgenic models without the risks of systemic exogenous administration. Early-phase clinical trials for related applications (IGF-1 gene therapy for heart failure) have been conducted, providing safety and feasibility data.
Biomarker-guided therapy. Identifying biomarkers that predict individual risk from IGF-1 elevation (genetic polymorphisms in IGF-1R, PI3K, PTEN, and other pathway components; baseline IGFBP profiles; cancer susceptibility markers) could enable personalized risk assessment that goes beyond the population-level hazard ratios from epidemiological studies. This precision medicine approach could eventually identify individuals for whom IGF-1-based therapy carries acceptable risk and those for whom it's contraindicated.
Combination approaches. Research into combining low-dose IGF-1 with other anabolic agents that work through different pathways (myostatin inhibitors, beta-2 agonists, selective androgen receptor modulators) could potentially achieve muscle growth comparable to high-dose IGF-1 monotherapy while reducing the IGF-1-specific risk exposure. This "combination, lower-dose" strategy parallels the approach used in modern cancer chemotherapy and may be applicable to anabolic pharmacology.
These research directions suggest that the current approach of injecting systemic LR3 may eventually be superseded by more sophisticated, targeted, and safer methods of harnessing IGF-1 biology for muscle growth and tissue repair. For those interested in staying current with developments in peptide science, the biohacking hub provides regularly updated coverage of emerging compounds and delivery technologies.
Summary and Clinical Perspective on Safety
Taking all safety considerations together, the risk profile of IGF-1 LR3 can be summarized as follows: acute risks are manageable with proper protocols (meal timing, glucose monitoring, appropriate dosing), but chronic risks - particularly the relationship between sustained IGF-1R activation and cancer susceptibility - represent a fundamental biological concern that cannot be mitigated through dosing strategy alone. The compound is not FDA-approved, has never been evaluated in human clinical trials, and creates a pharmacological state (sustained, unregulated IGF-1R activation) that has no safe analogue in normal human physiology.
For individuals who, after reviewing this evidence, decide to proceed with IGF-1 LR3 under medical supervision, the following principles should guide decision-making: use the lowest effective dose for the shortest feasible duration; monitor blood glucose continuously during initial dosing; obtain comprehensive baseline bloodwork including IGF-1 levels, metabolic panel, and age-appropriate cancer screening; never use LR3 in the presence of any active or suspected malignancy; avoid combining with insulin; and maintain a regular schedule of follow-up monitoring throughout use and during the post-cycle recovery period.
The free assessment at FormBlends can help determine whether peptide therapy is appropriate for your specific goals and health status, and the science and research section provides additional background on the evidence supporting various peptide therapies.
Frequently Asked Questions
What is IGF-1 LR3 and how does it work?
IGF-1 LR3 (Long R3 Insulin-Like Growth Factor-1) is a synthetic 83-amino-acid protein analog of human IGF-1. It works by binding to and activating the type 1 IGF receptor (IGF-1R), triggering two major downstream signaling pathways: PI3K/Akt/mTOR (which stimulates protein synthesis and inhibits protein degradation) and MAPK/ERK (which promotes cell proliferation and differentiation). The "LR3" designation reflects its two key modifications: a 13-amino-acid N-terminal extension ("Long") and an arginine substitution at position 3 ("R3"). These changes reduce binding to IGF binding proteins by roughly 100-fold, resulting in a half-life of 20-30 hours compared to native IGF-1's approximately 12 minutes in its free form. This makes LR3 approximately 2-3 times more potent than unmodified IGF-1 per unit dose.
How does IGF-1 LR3 differ from natural IGF-1?
Natural IGF-1 is a 70-amino-acid hormone produced primarily by the liver in response to growth hormone stimulation. In circulation, approximately 98-99% of native IGF-1 is bound to insulin-like growth factor binding proteins (IGFBPs), leaving only 1-2% in the free, biologically active form. IGF-1 LR3 differs in three key respects: it contains 13 additional amino acids at the N-terminus, has an arginine replacing glutamic acid at position 3, and consequently has dramatically reduced IGFBP affinity. The practical result is that LR3 circulates predominantly in free form for 20-30 hours, providing sustained, unregulated IGF-1 receptor activation. Native IGF-1's activity is tightly controlled by the IGFBP system, which regulates when, where, and how much IGF-1 reaches target receptors. LR3 bypasses this regulatory layer entirely.
What are the potential effects of IGF-1 LR3 on muscle growth?
IGF-1 LR3 promotes muscle growth through two complementary mechanisms. First, it stimulates muscle fiber hypertrophy by activating the PI3K/Akt/mTOR pathway, which increases protein synthesis rates by 20-40% while simultaneously suppressing protein degradation through FoxO transcription factor inhibition. Second, it activates satellite cells (muscle stem cells) through the MAPK/ERK pathway, promoting their proliferation and fusion with existing muscle fibers to provide additional myonuclei. Research using gamma irradiation to ablate satellite cells showed that approximately 50% of IGF-1-induced hypertrophy depends on satellite cell contribution. Transgenic mice with muscle-specific IGF-1 overexpression maintained youthful muscle mass into old age. However, no controlled human trials have been conducted specifically with LR3 for muscle growth.
What are the risks of IGF-1 LR3?
IGF-1 LR3 carries several categories of risk. Acutely, hypoglycemia is the most common and potentially dangerous side effect, reported in 42% of subjects receiving IGF-1 therapy in clinical trials, including cases of seizures and loss of consciousness. Fluid retention, headache, joint pain, and injection site reactions are also common. Chronically, the most serious concern is increased cancer risk. UK Biobank data from over 300,000 participants associates higher IGF-1 levels with increased breast cancer (HR=1.10), prostate cancer (HR=1.09), and colorectal cancer (HR=1.07) risk. The same PI3K/Akt/mTOR pathway that drives muscle growth also promotes cancer cell survival and proliferation. Organ enlargement, insulin resistance, cardiomyopathy, and suppression of the endogenous GH-IGF-1 axis are additional long-term concerns.
How does IGF-1 LR3 compare to IGF-1 DES?
IGF-1 LR3 and IGF-1 DES represent opposite engineering strategies for modifying IGF-1. LR3 adds amino acids (83 total) and has a 20-30 hour half-life, making it a systemic, long-acting compound. DES removes the first three amino acids (67 total) and has only a 20-30 minute half-life, making it a localized, short-acting compound. DES compensates for its brief duration with approximately 10-fold higher receptor binding affinity than native IGF-1. In practical terms, LR3 provides sustained whole-body anabolic signaling but carries higher cumulative risk from prolonged receptor activation. DES delivers intense but brief local stimulation when injected intramuscularly, potentially offering more site-specific effects with lower systemic exposure. The choice between them depends on whether the research goal is systemic versus localized IGF-1 receptor activation.
Is IGF-1 LR3 FDA-approved?
No, IGF-1 LR3 is not FDA-approved for any indication. The only FDA-approved IGF-1 product is mecasermin (brand name Increlex), which contains unmodified recombinant human IGF-1 and is approved specifically for long-term treatment of growth failure in pediatric patients with severe primary IGF-1 deficiency or GH gene deletion with neutralizing GH antibodies. Mecasermin was approved by the FDA in 2005 and by the European Medicines Agency in 2007. IGF-1 LR3 was originally developed as a cell culture supplement for biopharmaceutical production, not for therapeutic use. Its use in humans remains experimental and carries regulatory, legal, and safety implications that vary by jurisdiction. Products marketed as IGF-1 LR3 for human use are not manufactured under pharmaceutical GMP standards.
What is the typical dosing protocol for IGF-1 LR3?
Commonly referenced IGF-1 LR3 protocols use 20-60 mcg per day administered subcutaneously, with beginners typically starting at 20-30 mcg daily to assess tolerance. Cycles generally last 4-6 weeks, followed by an equal or longer off-period to allow endogenous GH-IGF-1 axis recovery. Injection timing is critical for safety: doses should be administered with or immediately before a meal containing both carbohydrates and protein to mitigate hypoglycemia risk. Post-workout injection timing is common, capitalizing on increased IGF-1 receptor sensitivity in exercised muscles. Doses above 50-60 mcg/day lack research support and substantially increase adverse effect risk. These protocols are derived from community experience and extrapolation from mecasermin clinical data, not from controlled clinical trials with LR3 itself.
Can IGF-1 LR3 cause cancer?
The relationship between IGF-1 signaling and cancer risk is well-established in the epidemiological literature. Higher circulating IGF-1 levels are consistently associated with modest but statistically significant increases in breast, prostate, and colorectal cancer risk across multiple large studies. The mechanistic basis involves the PI3K/Akt/mTOR pathway, which is one of the most commonly hyperactivated pathways in human cancers. IGF-1 receptor activation promotes cell survival, inhibits apoptosis, and stimulates cell proliferation - effects that can promote the growth of existing precancerous or cancerous cells. IGF-1 LR3 has never been specifically studied for cancer causation in humans, but its sustained, unregulated activation of the same receptor and pathways implicated in the epidemiological data provides a clear theoretical basis for concern. Individuals with active malignancy or cancer history should not use IGF-1 LR3.
What monitoring is needed when using IGF-1 LR3?
Comprehensive monitoring under medical supervision is essential for anyone using IGF-1 LR3. At minimum, this should include frequent blood glucose monitoring (a continuous glucose monitor is ideal, especially during initial dosing and overnight), baseline and periodic serum IGF-1 levels to assess degree of elevation, HbA1c before and after each cycle, complete metabolic panels to assess liver and kidney function, and adherence to age-appropriate cancer screening guidelines (PSA testing, mammography, colonoscopy per standard recommendations). A baseline echocardiogram is advisable for individuals with cardiovascular risk factors. Symptoms requiring immediate medical attention include severe hypoglycemia (confusion, seizures, loss of consciousness), persistent severe headache or visual changes (possible intracranial pressure elevation), and significant peripheral edema or shortness of breath.
How should IGF-1 LR3 be stored and reconstituted?
IGF-1 LR3 is typically supplied as a lyophilized powder in 1 mg vials. Unreconstituted powder should be stored at -20C (freezer) for long-term stability or 2-8C (refrigerator) for shorter periods. Reconstitution uses bacteriostatic water (containing 0.9% benzyl alcohol preservative), added slowly down the vial wall to avoid direct impact on the powder. Gentle swirling rather than shaking is recommended to prevent protein denaturation. A 1 mg vial reconstituted with 1 mL yields 1,000 mcg/mL; using 2 mL yields 500 mcg/mL. Reconstituted solution must be refrigerated at 2-8C and used within approximately 4 weeks with bacteriostatic water, or within 24-48 hours if reconstituted with plain sterile water. Never freeze reconstituted solution. Protect from light and heat exposure throughout storage and handling.
Additional Frequently Asked Questions
Can IGF-1 LR3 be used for injury recovery?
IGF-1 promotes tissue repair through multiple mechanisms: it stimulates fibroblast proliferation and collagen synthesis, enhances satellite cell activation for muscle fiber repair, promotes angiogenesis (new blood vessel formation) at injury sites, and protects cells from apoptosis during inflammatory phases. These properties make IGF-1 theoretically beneficial for muscle strain recovery, surgical recovery, and possibly tendon or ligament healing. However, clinical evidence specifically supporting IGF-1 LR3 for injury recovery in humans is absent. The systemic administration route means that the growth factor effects are not concentrated at the injury site, reducing the potential therapeutic benefit relative to the systemic risk exposure. Local injection at an injury site is sometimes discussed as an alternative, but this approach is unsupported by clinical data and carries risks of localized tissue effects. Peptides like BPC-157 and TB-500 have more specific tissue repair evidence and may be more appropriate for recovery-focused applications.
Does IGF-1 LR3 affect body fat?
IGF-1's effects on body fat are complex and context-dependent. In acute settings, IGF-1 has insulin-like effects that promote glucose uptake and lipogenesis (fat storage), particularly in the presence of elevated insulin. This contrasts with growth hormone, which has strong lipolytic (fat-burning) effects through direct activation of hormone-sensitive lipase in adipocytes. Over longer periods, increased lean muscle mass from IGF-1 signaling raises basal metabolic rate, which could indirectly support fat loss through increased daily energy expenditure. However, the net effect on body composition depends heavily on diet, exercise, and the specific dosing protocol. Some anecdotal reports describe mild body fat reduction during LR3 cycles, likely attributable to increased metabolic rate from muscle growth rather than direct lipolytic effects. For primary fat loss goals, GLP-1 receptor agonists like semaglutide or GH-based approaches have far stronger evidence.
What happens when you stop taking IGF-1 LR3?
Discontinuing IGF-1 LR3 triggers several physiological responses. First, the exogenous IGF-1R activation ceases over 24-48 hours as the remaining LR3 is cleared from circulation. Second, the suppressed endogenous GH-IGF-1 axis requires 2-4 weeks to recover full function, during which endogenous IGF-1 levels remain below normal baseline. This creates a transient state of relative IGF-1 deficiency that may manifest as increased muscle protein breakdown, reduced exercise recovery, fatigue, and potentially accelerated muscle loss. Third, the anabolic signaling environment shifts from net protein accretion back toward neutral or catabolic balance. Muscle gains made during the LR3 cycle can be partially preserved through continued resistance training and adequate protein intake, but some regression is expected, particularly in individuals who were at or above their natural genetic ceiling for muscle mass during the cycle. The magnitude of post-cycle muscle retention is highly individual and depends on training, nutrition, and whether the endogenous axis recovery is supported with appropriate strategies.
Is IGF-1 LR3 detectable in drug testing?
Yes, IGF-1 LR3 is detectable through modern anti-doping testing methods. The World Anti-Doping Agency (WADA) classifies all IGF-1 analogs as prohibited substances under Category S2 (Peptide Hormones, Growth Factors, Related Substances and Mimetics). Detection methods include immunoassay-based screening that can detect elevated total IGF-1 levels, and more specific mass spectrometry-based techniques that can identify the unique molecular signature of the LR3 variant (specifically the N-terminal extension and Glu3Arg substitution) distinct from endogenous IGF-1. The detection window depends on the dose and duration of use, but given LR3's 20-30 hour half-life, traces may be detectable for several days after the last injection through direct detection methods. Indirect markers (elevated IGF-1 with suppressed GH, altered IGFBP profiles) may persist longer. Athletes subject to WADA testing should be aware that positive tests result in suspension and potential career consequences.
Can IGF-1 LR3 be combined with peptides like BPC-157 or TB-500?
Combining IGF-1 LR3 with tissue repair peptides like BPC-157 or TB-500 is a relatively common practice in the peptide research community, though no formal clinical studies have examined these specific combinations. From a pharmacological perspective, these compounds work through largely independent receptor systems: IGF-1 LR3 activates the IGF-1R/PI3K/Akt/mTOR cascade; BPC-157 works through GH receptor upregulation, nitric oxide modulation, and VEGF-mediated angiogenesis; TB-500 acts through actin binding, cellular migration, and anti-inflammatory pathways. The minimal pharmacological overlap suggests a lower risk of dangerous complementary effects compared to combining LR3 with insulin or growth hormone. However, the absence of formal interaction studies means that unexpected effects cannot be ruled out. If combining these compounds, starting each at a conservative dose and introducing them sequentially (rather than simultaneously) allows for clearer attribution of effects and side effects.
What is the difference between IGF-1 LR3 and growth hormone therapy?
Growth hormone (GH) and IGF-1 LR3 operate at different levels of the same biological axis but have important distinctions. GH is produced by the pituitary gland and acts on cells throughout the body, where it has both direct effects (lipolysis in adipocytes, glucose regulation in the liver) and indirect effects mediated through stimulating IGF-1 production (primarily in the liver). IGF-1 LR3 bypasses the GH step entirely, directly activating the IGF-1 receptor without the GH-specific direct effects. Key practical differences include: GH promotes fat burning while LR3 has insulin-like lipogenic effects; GH preserves physiological IGFBP regulation while LR3 bypasses it; GH has decades of post-market safety data across multiple approved indications while LR3 has none; GH therapy raises both GH and IGF-1 while LR3 raises IGF-1 receptor activation but suppresses endogenous GH; and GH maintains pulsatile signaling patterns while LR3 produces continuous receptor activation. For most clinical applications, GH or GH secretagogues offer a more favorable risk-benefit profile than direct IGF-1 administration.
Practical Decision Framework for Clinicians and Researchers
For healthcare providers considering IGF-1 LR3 as part of a clinical protocol, a structured decision-making framework can help organize the relevant factors:
Step 1: Assess the clinical objective. Is the goal muscle growth, tissue repair, anti-aging, or research? For each objective, consider whether IGF-1 LR3 offers advantages over safer alternatives with established evidence bases. For muscle growth, resistance training combined with adequate protein intake and possibly GH secretagogues represents a first-line approach with dramatically lower risk. For tissue repair, BPC-157/TB-500 blends offer targeted healing without the systemic growth factor risks. For anti-aging, NAD+, Epithalon, and SS-31 target aging mechanisms through pathways that don't carry oncogenic concerns.
Step 2: Evaluate patient-specific risk factors. Personal or family history of cancer (especially breast, prostate, or colorectal) substantially increases the risk profile. Metabolic conditions (diabetes, insulin resistance) complicate glucose management. Cardiovascular disease increases vulnerability to fluid retention and cardiac effects. Age affects both the potential benefit (greater in older adults with declining IGF-1) and the baseline cancer risk (higher in older adults).
Step 3: Establish monitoring infrastructure. Before prescribing or recommending IGF-1 LR3, ensure that the patient has access to blood glucose monitoring (preferably CGM), baseline laboratory assessment (IGF-1, metabolic panel, HbA1c, PSA/mammography as appropriate), and follow-up appointments for ongoing monitoring. Without this infrastructure, the acute safety risks (particularly hypoglycemia) are unacceptably high.
Step 4: Informed consent. Given the experimental nature of LR3 use, thorough informed consent is essential. Patients should understand that the compound is not FDA-approved, has no human clinical trial data, carries meaningful cancer risk implications based on epidemiological evidence, and has an unknown therapeutic index. Document this discussion thoroughly.
Step 5: Protocol design. Use the lowest dose expected to produce effects (start at 20-30 mcg/day), limit cycle duration (4-6 weeks maximum), mandate meal timing with every injection, schedule regular monitoring, and have a clear plan for managing adverse effects (hypoglycemia protocol, emergency glucagon availability). The dosing calculator can assist with protocol development.
Step 6: Re-evaluate regularly. After each cycle, assess whether the observed benefits justify the risks. If the response is minimal relative to the risk exposure, discontinuation and transition to safer alternatives is the appropriate course. If the response is meaningful and the patient is informed and committed, careful continuation with ongoing monitoring may be reasonable.
This framework emphasizes that IGF-1 LR3 should be considered a last-resort option in most clinical contexts, appropriate only when safer alternatives have been tried and found insufficient, and when the patient fully understands and accepts the risk profile. The free assessment can help patients and clinicians determine which peptide therapy approach best matches individual goals and risk tolerance.
References
- Musaro A, McCullagh K, Paul A, Houghton L, Dobrowolny G, Molinaro M, Barton ER, Sweeney HL, Rosenthal N. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nature Genetics. 2001;27(2):195-200. DOI: 10.1038/84839. PMID: 11175789.
- Adams GR, McCue SA. Localized infusion of IGF-I results in skeletal muscle hypertrophy in rats. Journal of Applied Physiology. 1998;84(5):1716-1722. DOI: 10.1152/jappl.1998.84.5.1716.
- Stitt TN, Drujan D, Clarke BA, Panaro F, Timofeyva Y, Kline WO, Gonzalez M, Yancopoulos GD, Glass DJ. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Molecular Cell. 2004;14(3):395-403. DOI: 10.1016/S1097-2765(04)00211-4. PMID: 15125842.
- Schiaffino S, Mammucari C. Regulation of skeletal muscle growth by the IGF1-Akt/PKB pathway: insights from genetic models. Skeletal Muscle. 2011;1(1):4. DOI: 10.1186/2044-5040-1-4. PMID: 21798082.
- Yoshida T, Delafontaine P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells. 2020;9(9):1970. DOI: 10.3390/cells9091970. PMID: 32858949.
- Murphy N, Knuppel A, Papadimitriou N, Martin RM, Tsilidis KK, Bravi F,"; Gunter MJ, et al. Circulating insulin-like growth factor-1 and risk of total and 19 site-specific cancers: cohort study analyses from the UK Biobank. Cancer Research. 2020;80(20):4588-4598. DOI: 10.1158/0008-5472.CAN-20-1281. PMID: 33067265.
- Tin Tin S, Reiter PL, et al. IGF-1 and risk of morbidity and mortality from cancer, cardiovascular diseases, and all causes in EPIC-Heidelberg. The Journal of Clinical Endocrinology & Metabolism. 2023;108(10):e1092-e1101. DOI: 10.1210/clinem/dgad188. PMID: 37066827.
- Lok F, Owens JA, Mundy L, Robinson JS, Owens PC. Long R3 insulin-like growth factor-I (IGF-I) infusion stimulates organ growth but reduces plasma IGF-I, IGF-II and IGF binding protein concentrations in the guinea pig. Journal of Endocrinology. 1996;148(1):77-87. PMID: 7561636.
- Chernausek SD, Backeljauw PF, Frane J, Kuntze J, Underwood LE. Long-term treatment with recombinant IGF-I in children with severe IGF-I deficiency due to growth hormone insensitivity. Journal of Clinical Endocrinology & Metabolism. 2007;92(3):902-910. DOI: 10.1210/jc.2006-1610. PMID: 17192294.
- Rosenblatt JD, Parry DJ. Gamma irradiation prevents compensatory hypertrophy of overloaded mouse extensor digitorum longus muscle. Journal of Applied Physiology. 1992;73(6):2538-2543. DOI: 10.1152/jappl.1992.73.6.2538.
- Jacquemin V, Furling D, Bigot A, Butler-Browne GS, Mouly V. IGF-1 induces human myotube hypertrophy by increasing cell recruitment. Experimental Cell Research. 2004;299(1):148-158. DOI: 10.1016/j.yexcr.2004.05.023.
- Bach MA, Bondy CA. Anatomy of the pituitary insulin-like growth factor system. Endocrinology. 1992;131(6):2588-2594. DOI: 10.1210/endo.131.6.1446598.
- Baxter RC. Signaling Pathways of the Insulin-like Growth Factor Binding Proteins. Endocrine Reviews. 2023;44(5):753-778. DOI: 10.1210/endrev/bnad008. PMID: 36974717.
- Chitnis MM, Yuen JS, Protheroe AS, Pollak M, Macaulay VM. The type 1 insulin-like growth factor receptor pathway. Clinical Cancer Research. 2008;14(20):6364-6370. DOI: 10.1158/1078-0432.CCR-07-4879.
- Holt RI, Sonksen PH. Growth hormone, IGF-I and insulin and their abuse in sport. British Journal of Pharmacology. 2008;154(3):542-556. DOI: 10.1038/bjp.2008.99. PMID: 18376417.
- Guevara-Aguirre J, Balasubramanian P, Guevara-Aguirre M, Wei M, Madia F, Cheng CW, Hwang D, Martin-Montalvo A, Saavedra J, Ingles S, de Cabo R, Cohen P, Longo VD. Growth hormone receptor deficiency is associated with a major reduction in pro-aging signaling, cancer, and diabetes in humans. Science Translational Medicine. 2011;3(70):70ra13. DOI: 10.1126/scitranslmed.3001845. PMID: 21325617.
- Frystyk J. Free insulin-like growth factors - measurements and relationships to growth hormone secretion and glucose homeostasis. Growth Hormone & IGF Research. 2004;14(5):337-375. DOI: 10.1016/j.ghir.2004.06.001.
- Colao A, Ferone D, Marzullo P, Lombardi G. Systemic complications of acromegaly: epidemiology, pathogenesis, and management. Endocrine Reviews. 2004;25(1):102-152. DOI: 10.1210/er.2002-0022.
- Musaro A, Giacinti C, Borsellino G, Dobrowolny G, Pelosi L, Cairns L, Ottolenghi S, Bernardi G, Cossu G, Battistini L, Molinaro M, Rosenthal N. Paracrine effects of IGF-1 overexpression on the functional decline due to skeletal muscle disuse: molecular and functional evaluation in hindlimb unloaded MLC/mIgf-1 transgenic mice. PLoS One. 2013;8(5):e65167. DOI: 10.1371/journal.pone.0065167.
- Rommel C, Bodine SC, Clarke BA, Rossman R, Nunez L, Stitt TN, Yancopoulos GD, Glass DJ. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nature Cell Biology. 2001;3(11):1009-1013. DOI: 10.1038/ncb1101-1009.
- Werner H, LeRoith D. The IGF1 signaling pathway: from basic concepts to therapeutic opportunities. International Journal of Molecular Sciences. 2023;24(19):14882. DOI: 10.3390/ijms241914882. PMID: 37834333.
- Clemmons DR. Role of IGF binding proteins in regulating IGF responses to changes in metabolism. Journal of Molecular Endocrinology. 2018;61(1):T139-T169. DOI: 10.1530/JME-18-0016.
- Renehan AG, Zwahlen M, Minder C, O'Dwyer ST, Shalet SM, Egger M. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysis. The Lancet. 2004;363(9418):1346-1353. DOI: 10.1016/S0140-6736(04)16044-3.
- Giustina A, Chanson P, Kleinberg D, Bronstein MD, Clemmons DR, Klibanski A, van der Lely AJ, Strasburger CJ, Lamberts SW, Ho KK, Casanueva FF, Melmed S. Expert consensus document: A consensus on the medical treatment of acromegaly. Nature Reviews Endocrinology. 2014;10(4):243-248. DOI: 10.1038/nrendo.2014.21.
- Francis GL, Ross M, Ballard FJ, Milner SJ, Senn C, McNeil KA, Wallace JC, King R, Wells JR. Novel recombinant fusion protein analogues of insulin-like growth factor (IGF)-I indicate the relative importance of IGF-binding protein and receptor binding for enhanced biological potency. Journal of Molecular Endocrinology. 1992;8(3):213-223. DOI: 10.1677/jme.0.0080213.
- Tomas FM, Knowles SE, Owens PC, Chandler CS, Francis GL, Read LC, Ballard FJ. Insulin-like growth factor-I (IGF-I) and especially IGF-I variants are anabolic in dexamethasone-treated rats. Biochemical Journal. 1993;292(Pt 3):857-862. DOI: 10.1042/bj2920857.
- Petrella JK, Kim JS, Mayhew DL, Cross JM, Bamman MM. Potent myofiber hypertrophy during resistance training in humans is associated with satellite cell-mediated myonuclear addition: a cluster analysis. Journal of Applied Physiology. 2008;104(6):1736-1742. DOI: 10.1152/japplphysiol.01215.2007.
- Philippou A, Maridaki M, Halapas A, Koutsilieris M. The role of the insulin-like growth factor 1 (IGF-1) in skeletal muscle physiology. In Vivo. 2007;21(1):45-54. PMID: 17354613.
- Firth SM, Baxter RC. Cellular actions of the insulin-like growth factor binding proteins. Endocrine Reviews. 2002;23(6):824-854. DOI: 10.1210/er.2001-0033.
- Yakar S, Liu JL, Stannard B, Butler A, Accili D, Sauer B, LeRoith D. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proceedings of the National Academy of Sciences. 1999;96(13):7324-7329. DOI: 10.1073/pnas.96.13.7324.
- Melmed S, Colao A, Barkan A, Molitch M, Grossman AB, Kleinberg D, Clemmons D, Chanson P, Laws E, Schlechte J, Vance ML, Ho K, Giustina A. Guidelines for acromegaly management: an update. Journal of Clinical Endocrinology & Metabolism. 2009;94(5):1509-1517. DOI: 10.1210/jc.2008-2421. PMID: 19208732.
- Guler HP, Zapf J, Froesch ER. Short-term metabolic effects of recombinant human insulin-like growth factor I in healthy adults. The New England Journal of Medicine. 1987;317(3):137-140. DOI: 10.1056/NEJM198707163170303.
- Savage MO, Camacho-Hubner C, David A, Metherell LA, Hwa V, Rosenfeld RG, Clark AJL. Idiopathic short stature: will genetics influence the choice between GH and IGF-I therapy? European Journal of Endocrinology. 2007;157(Suppl 1):S33-S37. DOI: 10.1530/EJE-07-0292.
- Federici M, Porzio O, Zucaro L, Fusco A, Borboni P, Lauro D, Sesti G. Distribution of insulin/insulin-like growth factor-I hybrid receptors in human tissues. Molecular and Cellular Endocrinology. 1997;129(2):121-126. DOI: 10.1016/S0303-7207(97)00042-6.
- Dey BR, Furlanetto RW, Bhatt H, Bhargava MM, Bhatt BA, Bhatt DL. Interaction of human suppressor of cytokine signaling (SOCS)-2 with the insulin-like growth factor-I receptor. Journal of Biological Chemistry. 2000;275(46):35553-35557. DOI: 10.1074/jbc.M006535200.