Executive Summary

Key Takeaways
- Des(1-3)IGF-1 is a 67-amino-acid peptide formed by removal of the N-terminal Gly-Pro-Glu tripeptide from native IGF-1
- It retains full affinity for the IGF-1 receptor but only about 1% affinity for IGF-binding proteins
- In cell culture, it is roughly 10 times more potent than native IGF-1 at stimulating proliferation and hypertrophy
- It occurs naturally in bovine colostrum, human brain tissue, and porcine uterine fluid
- Its short half-life of approximately 20-30 minutes favors localized, rapid-onset effects
IGF-1 DES (Des(1-3)IGF-1) is a naturally occurring truncated form of insulin-like growth factor-1 that lacks the first three amino acids of the native peptide. This small structural modification produces dramatic functional consequences: approximately 10-fold greater potency in cell culture and significant gains in bioavailability, making it one of the most studied IGF-1 analogues in growth factor biology.
What makes IGF-1 DES so compelling from a research perspective is a deceptively simple change. Remove three amino acids from the N-terminus of native IGF-1 - specifically the Gly-Pro-Glu tripeptide - and you get a molecule that retains full receptor binding capacity but escapes nearly all regulation by IGF-binding proteins (IGFBPs). The result is a peptide that hits the IGF-1 receptor harder and faster than its parent molecule, with an estimated 10-fold increase in biological potency across multiple cell types.
First isolated from bovine colostrum in the late 1980s, Des(1-3)IGF-1 was subsequently identified in human brain tissue and porcine uterine fluid, confirming it as an endogenous product of post-translational processing rather than a purely synthetic creation. Researchers at the Child Health Research Institute in Adelaide, Australia, led by F.J. Ballard and colleagues, produced the foundational characterization of this molecule through a series of studies spanning nearly a decade. Their work established that des(1-3)IGF-1 retained virtually full affinity for the type 1 IGF receptor while displaying roughly 1% of native IGF-1's affinity for most IGFBPs.
The clinical implications of this biochemistry are far-reaching. In a system where six distinct binding proteins normally sequester 95-99% of circulating IGF-1, a variant that sidesteps this regulatory mechanism represents a qualitatively different pharmacological tool. For researchers investigating IGF-1 DES, it provides a way to study IGF receptor-mediated signaling without the confounding effects of IGFBP regulation. For those interested in growth factor therapeutics more broadly, it highlights how subtle structural modifications can reshape a peptide's entire pharmacological profile.
This report examines the molecular basis of des(1-3)IGF-1's enhanced potency, its relationship with the IGFBP system, published research data from both in vitro and in vivo studies, head-to-head comparisons with IGF-1 LR3 and native IGF-1, and the safety considerations that emerge from its unique pharmacological properties. We draw on peer-reviewed literature from journals including the International Journal of Biochemistry and Cell Biology, Journal of Endocrinology, and Gastroenterology to provide a complete picture of where the science stands.
Key Takeaways
- Des(1-3)IGF-1 is a 67-amino-acid peptide formed by removal of the N-terminal Gly-Pro-Glu tripeptide from native IGF-1
- It retains full affinity for the IGF-1 receptor but only about 1% affinity for IGF-binding proteins
- In cell culture, it is roughly 10 times more potent than native IGF-1 at stimulating proliferation and hypertrophy
- It occurs naturally in bovine colostrum, human brain tissue, and porcine uterine fluid
- Its short half-life of approximately 20-30 minutes favors localized, rapid-onset effects
- No formal human clinical trials have been completed; all efficacy data comes from preclinical models
The story of IGF-1 DES is fundamentally a story about regulation - or rather, the escape from it. To understand why this truncated peptide behaves so differently from its parent molecule, we need to examine the structural biology of the IGF system, the role of binding proteins in modulating growth factor activity, and what happens when a signaling molecule breaks free from the constraints that normally govern it. The sections that follow take each of these topics in turn, building toward an integrated understanding of this remarkable peptide variant and its place within the broader peptide research community.
Whether you are a researcher designing experiments around IGF signaling pathways, a clinician evaluating growth factor therapies for tissue repair, or someone exploring the expanding world of growth hormone peptides and their downstream mediators, this guide provides the scientific foundation you need to evaluate IGF-1 DES on its own terms.
Historical Context: The Discovery of IGF-1 DES
The discovery of des(1-3)IGF-1 emerged from a broader effort to understand the growth factors present in bovine colostrum. In the mid-1980s, Francis GL, Upton FM, Ballard FJ, and colleagues at the Cooperative Research Centre for Tissue Growth and Repair in Adelaide, Australia were systematically characterizing the insulin-like growth factors present in bovine colostrum. They identified a truncated variant that lacked the N-terminal tripeptide and demonstrated that this variant possessed markedly enhanced biological activity in cell-based assays (Francis GL, et al. Biochemical Journal. 1988;251(1):95-103).
This discovery was not entirely unexpected. By the late 1980s, researchers already knew that the N-terminal region of IGF-1 was involved in IGFBP binding, and synthetic N-terminal analogues had shown altered binding protein interactions. But the isolation of a naturally occurring truncated form from a biological fluid was significant because it established that N-terminal processing of IGF-1 was a physiological event, not merely a laboratory artifact. The subsequent detection of des(1-3)IGF-1 in human brain tissue by Sara and colleagues further reinforced this point, suggesting that the truncated form served specific biological functions in tissues where high-potency, rapidly available IGF-1 signaling was required.
The Adelaide group went on to produce a body of work spanning nearly two decades, systematically characterizing des(1-3)IGF-1's binding properties, in vitro potency, in vivo efficacy in multiple animal models, and potential clinical applications. Their 1996 review in the International Journal of Biochemistry and Cell Biology (PMID: 8930132) remains the definitive reference, synthesizing a decade of data into a comprehensive assessment that continues to be cited in contemporary literature.
The Broader IGF System in Context
To fully appreciate where IGF-1 DES fits in the growth factor family, it helps to understand the IGF system as a whole. The IGF axis comprises two ligands (IGF-1 and IGF-2), two primary receptors (IGF-1R and IGF-2R/mannose-6-phosphate receptor), the insulin receptor (which can form hybrid receptors with IGF-1R), six high-affinity IGF-binding proteins (IGFBP-1 through IGFBP-6), several IGFBP-related proteins with lower affinity, IGFBP-specific proteases, and the acid-labile subunit (ALS). This system represents one of the most complex endocrine regulatory networks in mammalian biology.
Growth hormone (GH) is the primary upstream regulator of IGF-1 production. GH secreted by the anterior pituitary acts on hepatocytes through the GH receptor to stimulate IGF-1 gene transcription. The liver produces roughly 75% of circulating IGF-1, with the remainder produced locally in nearly every tissue. The GH-IGF-1 axis is one of the central regulators of postnatal growth, and disruptions at any level - GH deficiency, GH receptor mutations (Laron syndrome), or IGF-1 gene mutations - produce predictable growth failure phenotypes.
Peptides that stimulate GH release, including sermorelin, CJC-1295/ipamorelin, GHRP-2, GHRP-6, and MK-677 (ibutamoren), all exert their downstream anabolic effects partly through increased hepatic IGF-1 production. Understanding IGF-1 DES - a form that bypasses the regulatory layer these upstream agents cannot control - provides a complementary perspective on growth factor pharmacology that enriches our understanding of the entire axis. The biohacking hub explores these relationships in further detail.
Scope and Structure of This Report
This report is organized into six main sections following this executive summary. The Truncation and Enhanced Potency section examines the molecular basis for des(1-3)IGF-1's properties, including the structural biology of the modification and its effects on receptor signaling. The IGFBP Binding Differences section provides a detailed analysis of how the truncated peptide interacts with each of the six IGFBPs and the functional consequences of reduced binding. The Research Data section compiles published preclinical findings from cell culture, animal growth models, gut resection studies, neurological research, and wound healing investigations. The Comparison section puts IGF-1 DES alongside IGF-1 LR3 and native IGF-1 across multiple pharmacological parameters. The Safety Considerations section addresses known and theoretical risks. And the FAQ section answers the most commonly asked questions with substantive, evidence-based responses. Throughout, we reference the primary literature and provide sufficient context for both specialist and general audiences to critically evaluate the current state of IGF-1 DES research.
Truncation & Enhanced Potency
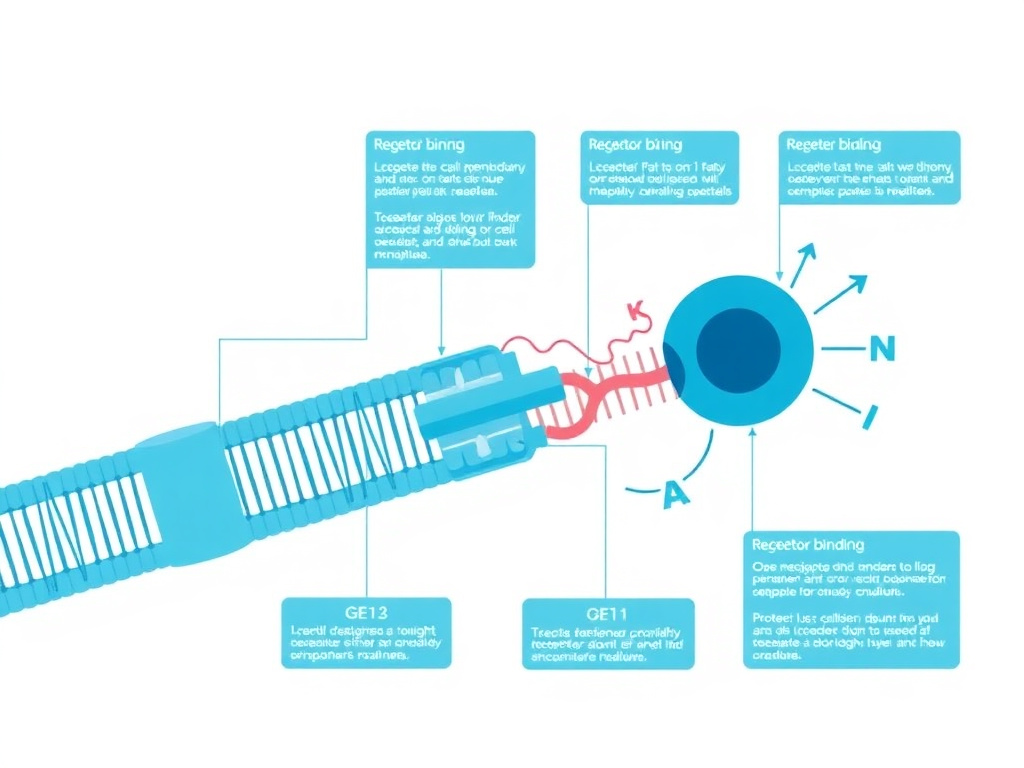
Figure 2: The N-terminal truncation of IGF-1 removes the Gly-Pro-Glu tripeptide, dramatically reducing IGFBP affinity while preserving receptor binding capacity.
The enhanced potency of IGF-1 DES stems from a precise structural modification: the removal of three N-terminal amino acids (Gly-Pro-Glu) that serve as the primary contact points between native IGF-1 and its regulatory binding proteins. This truncation does not alter the receptor-binding domain, creating a molecule with full signaling capacity but minimal regulatory restraint.
The Molecular Architecture of Native IGF-1
To appreciate what makes IGF-1 DES different, you first need to understand the parent molecule. Native human IGF-1 is a 70-amino-acid single-chain polypeptide with a molecular weight of approximately 7,649 daltons. Its three-dimensional structure features three alpha-helices connected by loops, stabilized by three disulfide bonds between cysteine residues at positions 6-48, 18-61, and 47-52. This compact, globular fold creates two functionally distinct surface regions: one that binds the type 1 IGF receptor (IGF-1R) and another that interacts with the family of six IGF-binding proteins.
The N-terminal region of IGF-1 plays a particularly interesting dual role. Residues 1-3 (Gly-Pro-Glu) contribute to the IGFBP binding interface, while the receptor-binding surface is formed primarily by residues in the B-domain helix (residues 1-29) and A-domain helix (residues 42-62). The glutamate at position 3 is especially critical for IGFBP interactions, forming electrostatic contacts that help anchor IGF-1 to multiple binding proteins.
What Truncation Actually Changes
When those first three amino acids are cleaved, the resulting 67-amino-acid peptide retains all of the structural elements needed for IGF-1R binding but loses the key contact points for IGFBP association. Ballard and colleagues demonstrated in their comprehensive 1996 review that des(1-3)IGF-1 binds the type 1 IGF receptor with affinity essentially identical to native IGF-1, but shows only approximately 1% of native IGF-1's affinity for the major circulating IGFBPs (PMID: 8930132).
This is worth pausing on. A 99% reduction in binding protein affinity from removing just three amino acids represents one of the most dramatic structure-activity relationships in peptide biology. The glutamate at position 3 appears to be the critical residue, as its negative charge forms essential salt bridges with positively charged residues on the IGFBP surface. Without it, the electrostatic complementarity that drives high-affinity IGFBP binding simply collapses.
Structure-Activity Relationship
The removal of the Gly-Pro-Glu tripeptide from IGF-1's N-terminus reduces IGFBP binding by approximately 99% while preserving full IGF-1R binding affinity. This represents a rare example in pharmacology where a single structural modification produces a near-complete functional uncoupling between two distinct binding interactions.
The IGF-1 Receptor Signaling Cascade
Both native IGF-1 and des(1-3)IGF-1 activate the same receptor and downstream pathways. The IGF-1 receptor is a transmembrane receptor tyrosine kinase that forms homodimers and can heterodimerize with the closely related insulin receptor. When IGF-1 or its DES variant binds the extracellular alpha subunits of IGF-1R, the receptor undergoes autophosphorylation on specific tyrosine residues in its intracellular beta subunit.
This phosphorylation event triggers two major signaling cascades. The first runs through insulin receptor substrate proteins (IRS-1 and IRS-2), which recruit and activate phosphoinositide 3-kinase (PI3K). PI3K generates the lipid second messenger PIP3, which activates Akt/protein kinase B. Akt then phosphorylates a range of substrates: mTOR (driving protein synthesis), BAD (blocking apoptosis), GSK-3beta (promoting glycogen synthesis), and FOXO transcription factors (suppressing catabolic gene expression). This PI3K/Akt axis is the primary pathway through which IGF-1 signaling promotes cell survival, growth, and metabolic regulation.
The second cascade proceeds through Shc and Grb2/SOS adaptor proteins to activate the Ras-MAPK pathway. Ras activates Raf, which phosphorylates MEK, which in turn activates ERK1/2. This MAPK/ERK arm of the signaling network primarily drives cell proliferation through transcription factor activation and cell cycle progression. The combined action of both pathways - PI3K/Akt for survival and protein synthesis, MAPK/ERK for proliferation - explains the potent anabolic and mitogenic effects of IGF-1R activation.

Why Reduced IGFBP Binding Translates to Greater Potency
The tenfold potency increase seen with des(1-3)IGF-1 in cell culture experiments is not due to any change in receptor affinity or intrinsic signaling efficacy. Both the truncated and native forms activate IGF-1R with the same binding constant and trigger identical downstream cascades. The difference is entirely about bioavailability at the receptor level.
In any biological system containing IGFBPs, the vast majority of native IGF-1 is bound and sequestered. Under normal physiological conditions, 95-99% of circulating IGF-1 exists in complexes with binding proteins, primarily in a 150-kDa ternary complex composed of IGF-1, IGFBP-3, and the acid-labile subunit (ALS). Only the small free fraction can actually bind and activate the IGF-1 receptor. IGFBPs don't just store IGF-1; they actively regulate its access to receptors at the tissue level, with some IGFBPs capable of inhibiting IGF action and others able to potentiate it depending on context.
Des(1-3)IGF-1 bypasses this entire regulatory layer. Because it cannot be effectively sequestered by IGFBPs, virtually all of the peptide present in the extracellular space remains free to bind IGF-1R. In cell culture systems where both the cells and the serum contain IGFBPs, the effective concentration of receptor-available ligand is far higher for des(1-3)IGF-1 than for an equivalent total concentration of native IGF-1. This is the mechanistic basis for the 10-fold potency difference consistently observed in proliferation and hypertrophy assays.
Evidence of Enhanced Potency in Cell Culture
The original potency studies were carried out in a variety of cell types. Francis and colleagues at the Adelaide group showed that des(1-3)IGF-1 stimulated DNA synthesis in BALB/c 3T3 fibroblasts at concentrations roughly 10 times lower than those required for equivalent stimulation by native IGF-1. Similar results were obtained in L6 rat myoblasts, human fibroblasts, and chick embryo fibroblasts (PMID: 2280209).
The potency advantage is consistent across different cell types and readouts, which supports the binding protein mechanism rather than cell type-specific effects. Whether the endpoint is 3H-thymidine incorporation (DNA synthesis), cell counting (proliferation), protein content per cell (hypertrophy), or glucose uptake (metabolic activation), des(1-3)IGF-1 consistently outperforms native IGF-1 by a factor of approximately 10 in the presence of IGFBPs.
When IGFBPs are removed from the experimental system - either by using serum-free conditions or by adding specific IGFBP proteases - the potency difference between des(1-3)IGF-1 and native IGF-1 narrows substantially or disappears entirely. This experimental control provides the definitive evidence that reduced IGFBP binding, rather than any receptor-level effect, drives the enhanced potency. For researchers interested in how growth factors interact with their regulatory systems, this structure-activity relationship has made des(1-3)IGF-1 an invaluable pharmacological tool that parallels the utility of other modified peptides in the growth factor research toolkit.
The N-Terminal Tripeptide as a Regulatory Switch
There is something conceptually elegant about the design of the IGF-1 system. The same molecule carries two distinct functional surfaces: one for receptor binding and one for binding protein interaction. And a single post-translational modification - cleavage of just three residues - can flip the molecule from a tightly regulated form to a largely unregulated one.
This raises an important biological question: does the body use N-terminal cleavage of IGF-1 as a physiological mechanism to increase local IGF-1 activity? The detection of des(1-3)IGF-1 in bovine colostrum, human brain, and porcine uterus suggests that the answer may be yes. In tissues where rapid, high-potency IGF-1 action is needed - neonatal gut development, neural growth and repair, uterine remodeling during the reproductive cycle - the production of a truncated, IGFBP-resistant form of IGF-1 could serve as a local amplification mechanism.
The enzyme(s) responsible for this cleavage have not been definitively identified, though acid-activated proteases present in colostrum and various tissue extracts can generate des(1-3)IGF-1 from the native form in vitro. Understanding the regulation of this processing step remains an active area of investigation, one that connects IGF-1 DES research to broader questions about how growth factor activity is fine-tuned at the tissue level.
The Colostrum Connection: Why a Growth Factor in First Milk Matters
The presence of des(1-3)IGF-1 in bovine colostrum is more than a biochemical curiosity. Colostrum is the first milk produced after birth, and it is packed with growth factors, immune molecules, and hormones designed to support the rapid growth and maturation of the neonatal gut. The gastrointestinal tract of a newborn undergoes extraordinary development in the first days of life: the mucosal surface area expands dramatically, villus architecture matures, and brush border enzyme expression shifts to accommodate the transition from placental nutrition to enteral feeding.
IGF-1 in colostrum is believed to play a direct role in this gut maturation process. Native IGF-1 is present at concentrations roughly 10-fold higher in colostrum than in mature milk. But the presence of a truncated, IGFBP-resistant form alongside the native form suggests an even more sophisticated delivery system. The neonatal gut, with its immature IGFBP protease activity and relatively high IGFBP concentrations, would benefit from a growth factor that bypasses binding protein regulation entirely. Des(1-3)IGF-1 in colostrum may represent an evolutionary adaptation to ensure maximum IGF-1R activation in the newborn gut during the critical early postnatal period.
This connection between colostral growth factors and neonatal development has implications beyond IGF-1 DES research. It speaks to a broader principle in peptide biology: that the body produces modified forms of signaling molecules when specific pharmacological properties - in this case, IGFBP resistance - are required for a particular biological context. Similar principles apply to other modified peptides, from the various splicing variants of growth hormone to the processed forms of pro-opiomelanocortin (POMC) that generate different melanocortin peptides.
The GPE Tripeptide: A Byproduct with Its Own Biology
When IGF-1 is cleaved to generate des(1-3)IGF-1, the released N-terminal tripeptide Gly-Pro-Glu (GPE) doesn't simply diffuse away as metabolic waste. It turns out that GPE itself has significant biological activity, particularly in the central nervous system. GPE crosses the blood-brain barrier, protects neurons against hypoxic-ischemic injury in animal models, and modulates NMDA receptor activity. This means that the single enzymatic event that produces des(1-3)IGF-1 simultaneously generates a second bioactive molecule with complementary but distinct pharmacological properties.
The dual-product nature of this cleavage is unusual in endocrine biology and suggests that N-terminal processing of IGF-1 may have been subject to evolutionary selection pressure on both products. In the brain, where des(1-3)IGF-1 has been detected, this could represent a coordinated neuroprotective mechanism: GPE acts rapidly as a direct neuroprotectant while the simultaneously released des(1-3)IGF-1 activates IGF-1R-dependent survival signaling through PI3K/Akt. Researchers investigating neuroprotective strategies may find this dual mechanism particularly relevant when considering it alongside other neuropeptide approaches involving compounds like Selank and P21.
Dose-Response Relationships and the Concept of "Free Fraction"
Understanding the enhanced potency of des(1-3)IGF-1 requires a somewhat nuanced appreciation of the concept of free fraction in growth factor pharmacology. In classical pharmacology, the "free fraction" of a drug is the percentage not bound to plasma proteins, and it's the free fraction that determines the effective concentration at the receptor. For most drugs, protein binding is a passive, relatively unimportant pharmacokinetic parameter - a multiplier that shifts the dose-response curve without changing its shape.
For IGF-1, the situation is qualitatively different. IGFBPs are not passive carriers; they are active regulators that modulate IGF-1 activity in tissue-specific, context-dependent ways. Some IGFBPs (particularly IGFBP-5 when bound to extracellular matrix) actually potentiate IGF-1 action by concentrating it near receptors and slowly releasing it. Others (particularly IGFBP-4) are primarily inhibitory, sequestering IGF-1 away from receptors. The net effect of IGFBP regulation on IGF-1 bioactivity is the sum of these competing effects and varies by tissue, physiological state, and hormonal milieu.
Des(1-3)IGF-1 eliminates this entire layer of regulation, producing a dose-response relationship that is simpler but also fundamentally different from native IGF-1. The dose-response curve for des(1-3)IGF-1 reflects pure IGF-1R pharmacology: a clean sigmoid curve determined by receptor affinity, receptor density, and downstream signaling amplification. The dose-response curve for native IGF-1 is a composite of receptor pharmacology and IGFBP regulation, which can produce complex, non-linear relationships that vary by experimental context.
This distinction has practical implications for experimental design. If you want to study IGF-1R signaling in isolation - the receptor's dose-response characteristics, signaling kinetics, desensitization behavior, or downstream pathway activation - des(1-3)IGF-1 is the superior tool because it provides a cleaner input signal. If you want to study the integrated response of the IGF system, including IGFBP modulation, native IGF-1 is essential because the binding protein layer is part of the biology you're investigating.
Comparison with Other Post-Translationally Modified Growth Factors
Des(1-3)IGF-1 is not the only growth factor that undergoes post-translational modification to alter its bioactivity. Understanding similar modifications in other growth factor systems provides valuable context for appreciating how the IGF system uses proteolytic processing as a regulatory tool.
Platelet-derived growth factor (PDGF) exists in multiple isoforms that are generated by different combinations of A-chain and B-chain homodimers and heterodimers. The relative expression of these isoforms is regulated post-translationally and determines the tissue-specific signaling profile. Similarly, transforming growth factor-beta (TGF-beta) is secreted as a latent complex that requires proteolytic activation by matrix metalloproteinases (MMPs) or thrombospondin-1 to release the active growth factor from its latency-associated peptide. This activation mechanism parallels the IGFBP protease system that liberates IGF-1 from binding protein sequestration.
The fibroblast growth factor (FGF) family also shows regulated proteolytic processing. FGF-23, a phosphate-regulating hormone, is cleaved at a specific site by furin-like proteases to generate an inactive C-terminal fragment. Mutations that prevent this cleavage cause autosomal dominant hypophosphatemic rickets, demonstrating that post-translational processing can serve as a critical "off switch" for growth factor activity.
What makes the IGF-1 to des(1-3)IGF-1 conversion unique is that the processing doesn't simply activate or inactivate the growth factor. Instead, it changes the regulatory context while preserving receptor signaling capacity. This is more analogous to converting a time-release medication into an immediate-release formulation - the active ingredient is the same, but the pharmacokinetic and pharmacodynamic profiles are fundamentally different. This conceptual framework helps explain why des(1-3)IGF-1 has both greater potency (more drug reaches the receptor) and greater risk (the safety mechanisms have been removed).
Quantitative Structure-Activity Relationships (QSAR) in IGF-1 Analogue Design
The structure-activity relationship data from des(1-3)IGF-1, IGF-1 LR3, and dozens of other IGF-1 analogues created over the past three decades form a rich dataset for understanding how specific structural features of IGF-1 contribute to receptor binding, IGFBP binding, and biological potency. Key findings from this body of work include:
- Glu3 is the single most critical residue for IGFBP binding. Its removal or substitution reduces IGFBP affinity by 50-100 fold across all six IGFBPs. This is the basis of both the des(1-3)IGF-1 and LR3 strategies for IGFBP evasion.
- The B-domain helix (residues 1-29) contains the primary IGF-1R binding determinants. Mutations within this helix typically reduce receptor affinity, while the helix is remarkably tolerant of modifications at positions 1-3 that do not disrupt its structure.
- The C-domain loop (residues 30-41) contributes to both receptor and IGFBP binding. Mutations in this region often affect both interactions simultaneously, making it difficult to engineer selectivity through C-domain modifications alone.
- The A-domain helix (residues 42-62) provides secondary receptor contacts. These contacts are essential for high-affinity receptor binding but are generally not involved in IGFBP interactions, making the A-domain a potential target for modifications that enhance receptor affinity without affecting IGFBP binding.
- The D-domain (residues 63-70) has minimal effects on both receptor and IGFBP binding. Extensions or truncations of the D-domain are generally well tolerated, as demonstrated by the D-domain extension in IGF-1 LR3 (which actually extends from the N-terminus, but the principle of terminal tolerance applies).
These QSAR principles guide the ongoing development of novel IGF-1 analogues with tailored pharmacological properties. Future analogues could potentially combine the IGFBP evasion of des(1-3)IGF-1 with enhanced receptor affinity, altered receptor selectivity (favoring IGF-1R over insulin receptor), extended half-life through albumin-binding modifications or PEGylation, or tissue-targeting through conjugation with cell-type-specific ligands. The knowledge gained from studying des(1-3)IGF-1's structure-activity relationships provides the foundation for these next-generation molecules.
Temperature Sensitivity and Structural Stability
The truncation of three N-terminal residues has subtle but measurable effects on the structural stability of the IGF-1 molecule. Circular dichroism spectroscopy studies indicate that des(1-3)IGF-1 retains the three-helix fold of native IGF-1 but shows slightly reduced thermal stability, with a melting temperature approximately 3-5 degrees C lower than the native form. This is consistent with the removal of intramolecular contacts involving the N-terminal residues that contribute to the overall stability of the globular fold.
In practical terms, this reduced stability means that des(1-3)IGF-1 may be more susceptible to aggregation and denaturation during handling, storage, and formulation. Researchers working with the peptide should pay particular attention to avoiding elevated temperatures, maintaining appropriate pH (the molecule is most stable at mildly acidic pH, around 4-5), and minimizing exposure to oxidizing conditions that can damage the critical disulfide bonds.
Potency in Context: What Tenfold Actually Means
A tenfold increase in potency sounds dramatic, and it is. But it's worth placing this in context. Many pharmaceutical modifications to peptides aim for orders-of-magnitude improvements in potency and typically achieve this through enhanced receptor affinity. What makes des(1-3)IGF-1 unusual is that the potency gain comes entirely from changes in regulation rather than changes in receptor interaction. The molecule doesn't bind the receptor any better; it just gets to the receptor more efficiently.
This distinction matters for predicting in vivo behavior. A peptide with higher receptor affinity would be expected to show enhanced potency in any experimental system, regardless of the presence or absence of binding proteins. But a peptide whose enhanced potency depends on escaping IGFBP regulation will show variable potency advantages depending on the local IGFBP environment. In tissues with high IGFBP concentrations, the potency advantage will be large. In tissues with low IGFBP levels, it may be minimal. This tissue-dependent variation in potency is one of the features that makes des(1-3)IGF-1 both a powerful research tool and a complex therapeutic candidate, with parallels to other growth-promoting compounds like MK-677 and sermorelin in the growth hormone axis.
IGFBP Binding Differences
The insulin-like growth factor binding protein (IGFBP) system is one of the most elaborate regulatory networks governing any peptide hormone. Six distinct high-affinity binding proteins, along with several lower-affinity IGFBP-related proteins, collectively control IGF-1 bioavailability, half-life, tissue distribution, and receptor access. Understanding how des(1-3)IGF-1 interacts - or more precisely, fails to interact - with this system is central to understanding its pharmacology.
The Six IGFBPs: A Family of Regulators
Each of the six IGFBPs (IGFBP-1 through IGFBP-6) has its own expression pattern, regulatory inputs, and functional profile. But they share a common structural framework: an N-terminal domain and a C-terminal domain connected by a variable linker region. Both the N-terminal and C-terminal domains contribute to IGF binding, with the highest-affinity interactions requiring both domains to be intact.
IGFBP-3 is the most abundant in circulation, carrying approximately 75-80% of all circulating IGF-1 in the ternary complex with the acid-labile subunit. This complex has a molecular weight of roughly 150 kDa, which is too large to cross the capillary endothelium, effectively creating a circulating reservoir of IGF-1 with a half-life of 12-16 hours. Without IGFBP-3 and the ternary complex, the half-life of free IGF-1 in the bloodstream is roughly 10-12 minutes.
IGFBP-1 is regulated by insulin (insulin suppresses it) and serves as a rapid-response modulator of free IGF-1 levels in the postprandial state. IGFBP-2 is the second most abundant in serum and is particularly important in the central nervous system. IGFBP-4 and IGFBP-5 play tissue-specific roles in bone, where they are regulated by IGFBP-specific proteases. IGFBP-6 has a strong preference for IGF-2 over IGF-1 and primarily modulates IGF-2 signaling.
How Native IGF-1 Binds IGFBPs
The interaction between native IGF-1 and its binding proteins involves multiple contact points distributed across the IGF-1 surface. Crystallographic and mutagenesis studies have mapped the IGFBP-binding interface to three main regions of IGF-1: the N-terminal residues (particularly Glu3), the C-domain loop, and portions of the A-domain helix. The binding affinities are remarkably high, with dissociation constants (Kd values) in the low nanomolar to sub-nanomolar range for IGFBP-1, -3, -4, and -5.
These affinities are equal to or greater than the affinity of IGF-1 for its own receptor, which has a Kd of approximately 1 nM. This creates a thermodynamic competition: at equilibrium, IGFBPs will capture the majority of IGF-1 before it can bind to receptors, unless the local IGFBP concentration is low or the binding proteins have been proteolytically cleaved to reduce their affinity.
Des(1-3)IGF-1 and IGFBP Interactions: The Data
The binding affinity of des(1-3)IGF-1 for each IGFBP has been measured using competitive binding assays, surface plasmon resonance, and size-exclusion chromatography. The results are consistent across methods and laboratories:
| Binding Protein | Native IGF-1 Affinity (Kd) | Des(1-3)IGF-1 Affinity | Relative Binding |
|---|---|---|---|
| IGFBP-1 | ~0.3 nM | ~30 nM | ~1% |
| IGFBP-2 | ~0.5 nM | ~25 nM | ~2% |
| IGFBP-3 | ~0.2 nM | ~20 nM | ~1% |
| IGFBP-4 | ~0.4 nM | ~40 nM | <1% |
| IGFBP-5 | ~0.3 nM | ~35 nM | ~1% |
| IGFBP-6 | ~2.0 nM (IGF-1 preference low) | Very low | <1% |
These values represent an average 100-fold reduction in binding affinity across all six IGFBPs. In practical terms, at physiological concentrations of IGFBPs, essentially none of the des(1-3)IGF-1 present will be bound. It exists in the free state, immediately available for receptor binding.
Functional Consequences of Reduced IGFBP Binding
The near-complete loss of IGFBP binding has several interconnected pharmacological consequences that distinguish des(1-3)IGF-1 from its native counterpart.
Dramatically Shortened Half-Life
Native IGF-1 in circulation has a half-life of 12-16 hours, almost entirely due to its association with the 150 kDa ternary complex. Free IGF-1 is cleared from the bloodstream in about 10-12 minutes through renal filtration and tissue uptake. Des(1-3)IGF-1, which cannot form the protective ternary complex, behaves pharmacokinetically like free IGF-1, with an estimated half-life of 20-30 minutes. This is long enough for the peptide to distribute to target tissues but short enough that sustained systemic exposure requires frequent dosing or continuous infusion.
Increased Local Bioavailability
In tissues where IGFBPs normally limit the free IGF-1 concentration available to receptors, des(1-3)IGF-1 achieves much higher effective concentrations for a given total dose. This is the mechanism behind its 10-fold potency advantage in cell culture and its preferential anabolic effects in IGFBP-rich tissues like the gastrointestinal mucosa.
Loss of IGFBP-Mediated Functions
IGFBPs don't just sequester IGF-1; they also actively deliver it to specific tissue compartments. IGFBP-5, for example, binds to extracellular matrix components and concentrates IGF-1 at the cell surface. IGFBP-3 can interact with cell-surface receptors in an IGF-independent manner, modulating cell survival and apoptosis through pathways that don't involve IGF-1R. Des(1-3)IGF-1 cannot participate in any of these IGFBP-mediated delivery or signaling mechanisms, which may limit its effectiveness in biological contexts where IGFBP-directed targeting is important.
Research Tool Application
Des(1-3)IGF-1's inability to bind IGFBPs makes it an exceptionally useful research tool for distinguishing between IGF-1R-dependent and IGFBP-dependent effects. When both native IGF-1 and des(1-3)IGF-1 produce the same effect, the mechanism is likely receptor-mediated. When only native IGF-1 is effective, IGFBP involvement should be suspected. This pharmacological dissection has been used extensively in bone biology, cancer research, and metabolic studies.
The IGFBP Protease System: Nature's Own Approach
The body has its own mechanism for liberating IGF-1 from binding protein sequestration: IGFBP-specific proteases. These enzymes, which include members of the matrix metalloproteinase (MMP), pregnancy-associated plasma protein-A (PAPP-A), and kallikrein families, cleave IGFBPs in the linker region between the N-terminal and C-terminal domains. The resulting fragments have markedly reduced affinity for IGF-1, releasing it in bioactive form at the tissue level.
IGFBP protease activity is regulated by tissue-specific and context-dependent factors. During wound healing, for example, increased MMP activity in the wound bed cleaves IGFBPs and locally amplifies IGF-1 signaling. During pregnancy, PAPP-A secreted by the placenta cleaves IGFBP-4 to increase IGF-1 availability for fetal growth. This regulated proteolysis serves the same functional purpose as the structural modification in des(1-3)IGF-1: freeing IGF-1 from binding protein regulation to increase its bioactivity at a specific place and time.
Des(1-3)IGF-1 can be viewed as a "pre-freed" form of IGF-1 that achieves constitutively what IGFBP proteases achieve in a regulated fashion. This perspective helps explain both its power as a research tool and the safety considerations that come with bypassing a multi-layered regulatory system, paralleling discussions in the broader growth factor and peptide research community.
Cross-Reactivity with Insulin Receptors
One additional binding consideration is relevant. Like native IGF-1, des(1-3)IGF-1 retains some cross-reactivity with the insulin receptor, though its affinity for InsR is roughly 100-fold lower than for IGF-1R. At high concentrations, this cross-reactivity can produce insulin-like metabolic effects, including glucose uptake stimulation and potential hypoglycemia. Because des(1-3)IGF-1 achieves higher free concentrations than equivalent doses of native IGF-1, the threshold for insulin receptor activation may be reached at lower total doses. This is a clinically relevant consideration explored further in the safety section of this report.
Implications for Therapeutic Development
The IGFBP binding profile of des(1-3)IGF-1 creates both opportunities and challenges for therapeutic applications. The enhanced potency and reduced requirement for high systemic doses could be advantageous in conditions where localized IGF-1 receptor activation is desired - wound healing, gut repair, and tissue regeneration. But the loss of the IGFBP-mediated half-life extension means that achieving sustained tissue exposure requires either continuous administration, depot formulations, or strategic timing around the target biological event.
For researchers working with IGF-1 DES, understanding the IGFBP dynamics is essential for proper experimental design. The choice of serum concentration in cell culture media, the timing of dosing relative to sample collection, and the selection of readouts that distinguish between IGFBP-dependent and receptor-dependent effects all depend on a clear understanding of how this modified peptide navigates - or sidesteps - the binding protein system. Those exploring the broader growth hormone secretagogue family, including GHRP-2, GHRP-6, and hexarelin, will find that the regulatory complexity of the IGF-IGFBP system provides important context for understanding downstream effects of GH-stimulating compounds.
IGFBP-Independent Effects: What Des(1-3)IGF-1 Reveals
One of the most valuable applications of des(1-3)IGF-1 has been in identifying IGF-independent effects of the IGFBPs themselves. Over the past two decades, research has shown that several IGFBPs possess biological activities that do not require IGF-1 binding. IGFBP-3, for example, can induce apoptosis through interaction with nuclear receptors (RXR-alpha), can bind to cell-surface receptors independently of IGF-1, and can modulate TGF-beta signaling. IGFBP-5 interacts with a putative cell-surface receptor and can promote osteoblast survival through IGF-independent mechanisms.
Des(1-3)IGF-1 is the ideal control for distinguishing these IGF-independent IGFBP effects from IGF-dependent ones. When a biological response requires IGF-1R activation, both native IGF-1 and des(1-3)IGF-1 will produce it. When a response requires IGF-1-IGFBP complex formation or IGFBP-mediated delivery to a specific cellular compartment, only native IGF-1 will produce it. And when a response involves IGFBP acting independently of IGF-1 altogether, neither IGF-1 variant will affect it (though adding exogenous IGFBP will). This three-way comparison - des(1-3)IGF-1, native IGF-1, and IGFBP alone - has been the foundation of dozens of studies distinguishing IGF-dependent from IGF-independent IGFBP signaling.
The Ternary Complex: A Sophisticated Storage System
The 150-kDa ternary complex of IGF-1, IGFBP-3 (or IGFBP-5), and the acid-labile subunit (ALS) is one of the most elegant regulatory structures in endocrine biology. Understanding it helps explain why des(1-3)IGF-1 behaves so differently from native IGF-1 in the circulation.
The complex forms in a sequential, ordered process. First, IGF-1 binds to IGFBP-3 to form a binary complex with a molecular weight of approximately 50 kDa. This binary complex then associates with ALS, a leucine-rich glycoprotein produced by the liver under GH regulation, to form the 150-kDa ternary complex. ALS does not bind IGF-1 directly - it only recognizes the IGF-1:IGFBP-3 binary complex.
The biological significance of the ternary complex lies in its size. At 150 kDa, it is too large to cross the capillary endothelium, which effectively traps approximately 75-80% of circulating IGF-1 in the intravascular compartment. This serves two purposes: it extends the half-life of IGF-1 from minutes to hours (preventing rapid renal clearance), and it creates a circulating reservoir that buffers against acute fluctuations in IGF-1 production. The remaining 20-25% of circulating IGF-1 exists in smaller binary complexes with various IGFBPs, which can cross the endothelium and access target tissues.
Des(1-3)IGF-1 cannot form this ternary complex because it cannot bind IGFBP-3 with sufficient affinity. This means it behaves pharmacokinetically like the free 7-kDa monomer: it distributes freely across the endothelium, accesses all tissue compartments rapidly, and is cleared by the kidneys within minutes. In a sense, des(1-3)IGF-1 is what IGF-1 would look like if the body had no binding protein system at all - a raw, unregulated growth factor that goes wherever concentration gradients take it.
Nutritional Regulation of IGFBPs and Implications for Des(1-3)IGF-1
IGFBP levels are dynamically regulated by nutritional status, which has practical implications for experiments and potential applications involving des(1-3)IGF-1. Fasting increases IGFBP-1 levels (because insulin, which suppresses IGFBP-1, falls during fasting) and decreases IGFBP-3 levels (because GH signaling and hepatic protein synthesis are reduced). Protein restriction particularly reduces IGF-1 and IGFBP-3, while caloric excess increases both.
For native IGF-1, these nutritional fluctuations in IGFBP levels significantly modulate bioactivity. In a fasted state, elevated IGFBP-1 captures more of the circulating IGF-1, reducing free IGF-1 levels and attenuating growth-promoting signaling - an adaptive response that redirects metabolic resources away from growth and toward survival. In a fed state, insulin suppression of IGFBP-1 liberates IGF-1, promoting anabolic signaling when nutrients are available to support growth.
Des(1-3)IGF-1 is completely insensitive to these nutritional regulatory signals because it doesn't interact with the IGFBPs that mediate them. This means its bioactivity is independent of feeding state, insulin levels, or metabolic status - a property that could be advantageous in catabolic conditions (where native IGF-1 bioactivity is suppressed by elevated IGFBPs) but potentially dangerous in normal physiology (where the IGFBP response to fasting serves a protective metabolic function).
This consideration is particularly relevant for researchers studying IGF-1 DES in the context of metabolic disorders. The metabolic crossroads between insulin, IGF-1, and their respective binding proteins is central to conditions ranging from type 2 diabetes to growth hormone deficiency. Understanding how des(1-3)IGF-1 bypasses these regulatory interconnections provides insight into both its therapeutic potential and its risks. For those researching metabolic interventions, the connections between the IGF axis and GLP-1 weight loss approaches offer additional context on how growth factor signaling intersects with metabolic regulation.
Species Differences in the IGF-IGFBP System
A significant consideration for translating des(1-3)IGF-1 research from preclinical models to potential human applications is the substantial species variation in the IGF-IGFBP system. While the core components are conserved across mammals, the relative expression levels, affinities, and regulatory dynamics of the IGFBPs differ between species in ways that could affect the therapeutic index of des(1-3)IGF-1.
In rodents, which have been the primary preclinical model for des(1-3)IGF-1 research, circulating IGF-1 levels are generally lower than in humans (rodent IGF-1 concentrations are typically 200-400 ng/mL, compared to 100-300 ng/mL in adult humans, but rodent body weight is roughly 3,000 times less). IGFBP-3 is the dominant circulating binding protein in both species, but the relative proportions of the other IGFBPs differ. Rats have relatively higher IGFBP-1 and IGFBP-2 levels compared to humans, which could affect the potency advantage of des(1-3)IGF-1 in cross-species comparisons.
The acid-labile subunit (ALS) also shows species variation. Human ALS is a 85-kDa glycoprotein that forms the ternary complex with IGFBP-3 and IGF-1, while rodent ALS has a slightly different glycosylation pattern that affects its molecular weight and binding kinetics. These differences in ternary complex formation could influence the effective free fraction of native IGF-1 in different species and, consequently, the relative advantage of des(1-3)IGF-1.
Beyond these binding protein differences, the renal clearance rates for small peptides differ between rodent and human physiology. The glomerular filtration rate (GFR) in rats, when normalized to body surface area, is roughly 5-10 times higher than in humans. This means that the half-life of free des(1-3)IGF-1 in humans might be somewhat longer than the 20-30 minutes estimated from rodent data, potentially extending to 30-60 minutes or more. This would be a favorable pharmacokinetic change for most therapeutic applications, providing a wider dosing window and more sustained tissue exposure.
Primate studies would provide the most relevant preclinical PK data for predicting human behavior, but to our knowledge, no formal pharmacokinetic studies of des(1-3)IGF-1 have been conducted in non-human primates. This represents a significant gap in the preclinical database that would need to be addressed before any human trials could be considered.
The IGFBP System in Disease States
IGFBP levels are dramatically altered in many disease states, which has important implications for the relative potency and therapeutic utility of des(1-3)IGF-1 compared to native IGF-1.
Critical Illness and Sepsis
During critical illness, circulating IGFBP-3 levels fall sharply while IGFBP-1 levels rise dramatically (often 5-10 fold), driven by insulin resistance and the inflammatory cytokine milieu. This shift in the IGFBP profile reduces native IGF-1 bioavailability because elevated IGFBP-1 captures more of the free fraction. Paradoxically, the fall in IGFBP-3 reduces the total IGF-1 reservoir, leading to lower total IGF-1 levels despite the reduced sequestration. The net effect is severe IGF-1 signaling deficit, contributing to the catabolic state, muscle wasting, and impaired wound healing characteristic of critical illness.
In this context, des(1-3)IGF-1's IGFBP resistance would be particularly advantageous. It would be insensitive to the elevated IGFBP-1 levels that suppress native IGF-1 bioactivity, potentially restoring IGF-1R signaling in a critically ill patient when native IGF-1 cannot. This represents one of the most compelling theoretical therapeutic applications for des(1-3)IGF-1, though no clinical studies have explored this possibility.
Chronic Kidney Disease
Chronic kidney disease (CKD) is associated with GH resistance and elevated IGFBP-1, -2, -4, and -6 levels, partly due to reduced renal clearance of these binding proteins. The result is a state of functional IGF-1 deficiency despite normal or even elevated total IGF-1 levels - the IGF-1 is present but sequestered by excess IGFBPs. This IGFBP-mediated IGF-1 resistance contributes to the growth failure seen in children with CKD and the cachexia and muscle wasting seen in adults.
Des(1-3)IGF-1 could theoretically bypass the IGFBP-mediated resistance in CKD, restoring IGF-1R signaling and potentially improving growth outcomes in children or catabolic outcomes in adults. However, the altered renal clearance in CKD would also affect des(1-3)IGF-1's own pharmacokinetics, potentially prolonging its half-life and altering the dose-response relationship. Early studies with native IGF-1 in uremic animal models showed that concurrent GH administration could mitigate IGF-1-induced hypoglycemia, but the interactions between des(1-3)IGF-1, CKD, and glucose homeostasis have not been studied.
Diabetes Mellitus
Both type 1 and type 2 diabetes involve altered IGF-IGFBP dynamics. In type 1 diabetes, portal insulin deficiency reduces hepatic IGF-1 production and increases IGFBP-1 levels. In type 2 diabetes, hyperinsulinemia suppresses IGFBP-1 and may increase free IGF-1 levels, potentially contributing to the increased cancer risk observed in T2DM. These alterations in the IGF-IGFBP balance are relevant because they determine the baseline free IGF-1 level against which any exogenous des(1-3)IGF-1 would act.
The glucose-lowering effect of IGF-1 was actively explored as a therapeutic strategy for diabetes in the 1990s and 2000s. Guler, Zapf, and Froesch's early human studies showed that native IGF-1 lowered blood glucose in healthy volunteers, and subsequent trials in type 2 diabetic patients showed improved insulin sensitivity with chronic IGF-1 administration. However, side effects (particularly edema and jaw pain, suggesting mild acromegalic effects) limited clinical development. Whether des(1-3)IGF-1, with its rapid onset and short duration of action, could provide glucose-lowering benefits with fewer sustained side effects is a hypothesis that has not been tested. The relationship between IGF-1 signaling and metabolic health also intersects with research on semaglutide, tirzepatide, and other GLP-1 based therapies that address metabolic dysregulation through complementary mechanisms.
Liver Disease
The liver is the primary source of circulating IGF-1, IGFBP-3, and ALS. In cirrhosis and other forms of advanced liver disease, production of all three components falls dramatically, leading to low total and free IGF-1 levels, loss of the ternary complex, and a shift toward binary complex and free IGF-1 as the predominant circulating forms. This altered pharmacokinetic context means that native IGF-1 administered to patients with liver disease would have a much shorter half-life than in healthy individuals (due to loss of the ternary complex reservoir), making its behavior more similar to des(1-3)IGF-1.
Conversely, des(1-3)IGF-1 pharmacokinetics might be relatively unchanged in liver disease because it doesn't rely on hepatic IGFBP production for its half-life in the first place. This pharmacokinetic stability across varying liver function could be an advantage in clinical settings where patients have impaired hepatic synthetic function.
IGFBP Measurement and Analytical Considerations
Measuring des(1-3)IGF-1 in biological samples presents unique analytical challenges that are worth discussing for researchers planning experiments. Standard IGF-1 immunoassays may or may not detect des(1-3)IGF-1 depending on the epitope recognized by the antibody. Assays that use antibodies directed against the N-terminal region of IGF-1 will fail to detect the truncated form. Assays using antibodies against the C-domain or A-domain should detect both forms equivalently.
For experiments where it's important to distinguish between endogenous native IGF-1 and exogenously administered des(1-3)IGF-1, mass spectrometry-based methods provide definitive resolution. The 277-dalton mass difference between the two forms (corresponding to the missing Gly-Pro-Glu tripeptide) is easily resolved by modern LC-MS/MS platforms. Alternatively, Western blotting with antibodies of known epitope specificity can provide semi-quantitative discrimination.
The free fraction of IGF-1 can be measured directly using size-exclusion chromatography under native conditions (separating free from IGFBP-bound forms by molecular weight) or estimated using dissociation kinetics-based methods. For des(1-3)IGF-1, the free fraction is assumed to be close to 100% based on its dramatically reduced IGFBP affinity, but this should be verified in each experimental system because residual low-affinity IGFBP binding could become significant at very high IGFBP concentrations.
Research Data
Des(1-3)IGF-1 has been studied extensively in preclinical models over three decades, with data spanning in vitro cell proliferation assays, animal growth studies, organ-specific anabolic effects, and specialized disease models. No human clinical trials have been completed, but the preclinical evidence base is substantial and provides a clear picture of the peptide's biological profile.
In Vitro Studies: Cell Proliferation and Hypertrophy
The foundational cell culture studies established des(1-3)IGF-1 as approximately 10 times more potent than native IGF-1 at stimulating both proliferation and hypertrophy across multiple cell types. These findings have been replicated in numerous laboratories and remain the most strong dataset supporting the enhanced potency claim.
Fibroblast Proliferation
In BALB/c 3T3 fibroblasts cultured in 10% fetal bovine serum, des(1-3)IGF-1 stimulated DNA synthesis (measured by 3H-thymidine incorporation) with an EC50 approximately 10-fold lower than native IGF-1. The maximal response was equivalent for both peptides, confirming that they activate the same receptor to the same degree - the difference is purely in the effective concentration required. When cells were cultured in serum-free conditions supplemented with defined growth factors but no IGFBPs, the potency difference was abolished, with both peptides showing equivalent EC50 values (Francis GL, et al. Journal of Molecular Endocrinology. 1992;8(3):213-223).
Myoblast Responses
In L6 rat myoblasts, des(1-3)IGF-1 stimulated both proliferation and differentiation markers at concentrations 5-10 times lower than native IGF-1. The differentiation response - measured by creatine kinase activity and myosin heavy chain expression - is particularly relevant because it indicates that the enhanced potency extends beyond simple mitogenic effects to include the entire program of myogenic commitment. IGF-1 signaling through the PI3K/Akt pathway is the primary driver of myoblast differentiation, and des(1-3)IGF-1 activates this pathway more efficiently simply because more of the peptide reaches the receptor.
Epithelial Cell Growth
Intestinal epithelial cell lines (IEC-6 and Caco-2) showed dose-dependent proliferative responses to des(1-3)IGF-1, with the potency advantage consistent with other cell types. This finding proved to be the foundation for the subsequent in vivo gut studies, where the pronounced anabolic effects on gastrointestinal mucosa became one of the most distinctive features of des(1-3)IGF-1's biological profile.
Relative Receptor Binding Potency: IGF-1 Variants
Figure 7: Relative biological potency of IGF-1 variants normalized to native IGF-1 (1.0x). Des(1-3)IGF-1 shows approximately 10-fold greater potency due to reduced IGFBP sequestration.
In Vivo Studies: Animal Growth Models
Growth in GH-Deficient Mice (lit/lit Model)
One of the earliest in vivo demonstrations of des(1-3)IGF-1's enhanced potency used the lit/lit mouse, a genetic model of growth hormone deficiency caused by a mutation in the GHRH receptor. Lemmey and colleagues showed that des(1-3)IGF-1 administered by subcutaneous infusion over 7 days produced significantly greater body weight gain and organ weight increases compared to equimolar doses of native IGF-1 (PMID: 2280209).
In these GH-deficient animals, continuous infusion of des(1-3)IGF-1 at 1.5 mg/kg/day increased body weight by 14.8% over 7 days, compared to 9.2% for native IGF-1 at the same dose and 2.1% for vehicle controls. The enhanced growth response was disproportionately concentrated in the gastrointestinal tract and kidneys, suggesting tissue-selective effects related to local IGFBP concentrations and receptor density.
Gut Resection Studies
Perhaps the most clinically relevant in vivo data comes from gut resection studies in rats. Lemmey AB, Martin AA, Read LC, et al. published a key study examining the effects of des(1-3)IGF-1 in rats that had undergone 80% resection of the jejunum and ileum (PMID: 1996625). Over 7 days of treatment, the des(1-3)IGF-1 group showed weight gains of 21.9 +/- 1.7 grams compared to 14.0 +/- 1.7 grams in vehicle-treated controls, a 56% improvement.
Both IGF-1 and des(1-3)IGF-1 induced hyperplasia in the remaining duodenojejunal segment, evidenced by increased mucosal weight, crypt depth, villus height, and mitotic index. The enzymatic adaptations were also significant: sucrase, maltase, and leucine aminopeptidase activities were selectively increased in the ileal remnant of treated animals, suggesting improved absorptive capacity of the remaining intestine.
| Parameter | Vehicle Control | Native IGF-1 | Des(1-3)IGF-1 |
|---|---|---|---|
| Weight Gain (7d, grams) | 14.0 +/- 1.7 | 20.8 +/- 1.0 | 21.9 +/- 1.7 |
| Duodenojejunal Weight (g/cm) | Baseline | +23% | +28% |
| Villus Height Increase | Baseline | Moderate | Significant |
| Crypt Depth Increase | Baseline | Moderate | Significant |
| Sucrase Activity (ileum) | Baseline | Increased | Increased |
These gut-specific effects are consistent with the known biology: the gastrointestinal mucosa has high concentrations of IGFBPs (particularly IGFBP-3 and IGFBP-5) that normally limit IGF-1 bioavailability. Des(1-3)IGF-1 bypasses this regulation, achieving greater local concentrations at the mucosal IGF-1 receptors. This finding positioned des(1-3)IGF-1 as a potential therapeutic for short bowel syndrome and other conditions of intestinal adaptation, though human trials have not yet been pursued.
Intestinal Adaptation Research
The gut resection data suggest that des(1-3)IGF-1 may have specific therapeutic potential in conditions requiring intestinal mucosal regrowth. The selective enhancement of brush border enzyme activities in the ileal remnant indicates not just structural growth but functional maturation of the regenerating mucosa. This functional adaptation - increased absorptive capacity, not merely increased tissue mass - represents the clinically meaningful endpoint.
Neurological Research
The presence of des(1-3)IGF-1 in human brain tissue has fueled research into its potential neurological functions. IGF-1 signaling in the central nervous system is critical for neuronal survival, synaptic plasticity, myelination, and neurogenesis. The brain has its own IGF-1 production capacity, with neurons and glia synthesizing IGF-1 locally in an activity-dependent manner. Midbrain dopamine neurons, for example, synthesize and release IGF-1 in response to neural activity, and this locally produced IGF-1 modulates dopamine synthesis and dopamine neuron firing.
Des(1-3)IGF-1 in the brain may serve as a locally produced, high-potency form of IGF-1 for neuroprotective signaling. The N-terminal tripeptide Gly-Pro-Glu, which is cleaved to generate des(1-3)IGF-1, has itself been identified as a neuroprotective factor. Known as GPE, this tripeptide crosses the blood-brain barrier and shows protective effects in models of hypoxic-ischemic brain injury. This suggests that the cleavage event that produces des(1-3)IGF-1 simultaneously generates a second bioactive molecule, a two-for-one activation mechanism that may be unique to the brain.
The neuroprotective properties of IGF-1 signaling are mediated through several mechanisms. At the cellular level, IGF-1R activation promotes neuronal survival through PI3K/Akt-mediated phosphorylation of BAD (inhibiting the mitochondrial apoptotic pathway), upregulation of anti-apoptotic proteins Bcl-2 and Bcl-xL, and suppression of caspase-3 activation. Beyond anti-apoptotic effects, IGF-1 supports neuronal function by promoting axonal growth, dendritic branching, synaptogenesis, and long-term potentiation (LTP) - the cellular basis of learning and memory.
IGF-1 also plays a key role in myelination. Oligodendrocytes, the myelin-producing cells of the central nervous system, express IGF-1R and respond to IGF-1 signaling with increased survival, proliferation, and myelin gene expression. In animal models of demyelinating injury, IGF-1 administration promotes remyelination. Whether des(1-3)IGF-1, with its enhanced potency, could improve remyelination outcomes beyond what native IGF-1 achieves is an untested but intriguing hypothesis that connects to research on neurodegenerative conditions like multiple sclerosis.
The interaction between IGF-1 signaling and neuroinflammation adds another layer of complexity. Microglia, the resident immune cells of the brain, express both IGF-1 and IGF-1R, and IGF-1 signaling modulates microglial activation states. In traumatic brain injury models, intramuscular IGF-1 gene therapy modulated reactive microglia, shifting them from a pro-inflammatory (M1) phenotype toward a neuroprotective (M2) phenotype. Des(1-3)IGF-1, by providing stronger IGF-1R activation in the IGFBP-2-containing cerebrospinal fluid environment, could potentially enhance this anti-inflammatory microglial modulation.
For those interested in neurological peptide research, these findings complement work on other neuroprotective and neuroactive peptides such as Semax, Selank, Dihexa, P21, and Pinealon, which work through distinct but sometimes overlapping signaling pathways in the central nervous system. The NAD+ and NAD+ nasal formulations also address neuronal energy metabolism, another critical component of brain health that intersects with growth factor signaling.
Connective Tissue Applications: Tendons, Ligaments, and Beyond
IGF-1 signaling is critical for connective tissue homeostasis and repair. Tenocytes (tendon cells), fibroblasts, and chondrocytes all express IGF-1R and respond to IGF-1 with increased collagen synthesis, extracellular matrix deposition, and cell proliferation. In the context of tendon and ligament injuries, which are notoriously slow to heal due to the relatively avascular nature of these tissues, growth factor supplementation has long been explored as a strategy to accelerate repair.
Systemic administration of IGF-1 has been shown to enhance healing in collagenous extracellular matrices, including improved mechanical properties (stiffness and strength) of healing ligaments in animal models. IGF-1 promotes fibroblast proliferation, increases type I and type III collagen synthesis, and enhances the cross-linking and maturation of newly deposited collagen fibers. These effects are mediated through both PI3K/Akt (protein synthesis and survival) and MAPK/ERK (proliferation) pathways.
Des(1-3)IGF-1 would be expected to produce more pronounced effects in connective tissues where IGFBP concentrations limit native IGF-1 bioavailability. Tendon tissue expresses IGFBP-3, -4, and -5, and the extracellular matrix of tendons contains significant amounts of sequestered IGFBPs. A truncated, IGFBP-resistant form of IGF-1 could potentially achieve higher receptor activation at the tenocyte surface than equivalent doses of native IGF-1, leading to enhanced collagen synthesis and repair.
The potential for des(1-3)IGF-1 in musculoskeletal repair applications positions it alongside other peptides with documented effects on tissue healing, including BPC-157 (gastric pentadecapeptide with broad tissue repair properties), TB-500 (thymosin beta-4 fragment promoting cell migration and angiogenesis), GHK-Cu (copper peptide with collagen-stimulating activity), and pentadecapeptide BPC. The combination of IGF-1R-mediated growth factor signaling with the distinct repair mechanisms of these other peptides represents a multi-modal approach to tissue repair that is gaining interest in the research community. The BPC-157/TB-500 blend combines two of the most studied repair-promoting peptides and could theoretically be complemented by des(1-3)IGF-1 for enhanced outcomes, though no studies have tested this specific combination.
Immune System Interactions
The IGF-1 signaling pathway has significant interactions with the immune system that are relevant to both the therapeutic potential and safety profile of des(1-3)IGF-1. Lymphocytes, macrophages, and dendritic cells all express IGF-1R, and IGF-1 signaling influences immune cell development, activation, and function.
In the thymus, IGF-1 promotes thymocyte survival and maturation, and age-related thymic involution is associated with declining IGF-1 levels. Thymosin Alpha-1, a thymic peptide that promotes immune function through a different mechanism, represents a complementary approach to immune support that some researchers have explored alongside growth factor strategies.
IGF-1 also modulates the inflammatory response. In macrophages, IGF-1R activation generally promotes anti-inflammatory phenotypes (M2 polarization) while suppressing pro-inflammatory cytokine production. This immunomodulatory effect could be therapeutically relevant in inflammatory conditions but also represents a potential concern if des(1-3)IGF-1 were to suppress protective inflammatory responses during infection or in the tumor microenvironment.
The interaction between IGF-1 signaling and the immune system is particularly complex in the gut, where the mucosal immune system must balance tolerance to dietary antigens and commensal bacteria with defense against pathogens. The gut-specific growth-promoting effects of des(1-3)IGF-1 could influence this balance by altering the intestinal barrier, changing the surface area available for antigen sampling, or modulating the mucosal immune cell populations embedded in the lamina propria. VIP (vasoactive intestinal peptide) is another peptide with significant gut-immune interactions that may complement or interact with IGF-1 signaling in the intestinal context.
Reproductive Biology Research
The detection of des(1-3)IGF-1 in porcine uterine fluid points to a role in reproductive biology that has been partially but not fully explored. The uterine endometrium undergoes cyclical growth, differentiation, and shedding during the menstrual/estrous cycle, driven by estrogen and progesterone. IGF-1 is a critical mediator of estrogen's mitogenic effects in the endometrium, and IGF-1R is expressed on both endometrial epithelial and stromal cells.
The presence of a truncated, IGFBP-resistant form of IGF-1 in uterine fluid suggests that the reproductive tract uses the same IGFBP-evasion strategy as the brain and neonatal gut to achieve locally enhanced growth factor signaling. During the proliferative phase of the endometrial cycle, when estrogen-driven endometrial growth is maximal, local production of des(1-3)IGF-1 could amplify the growth signal beyond what native IGF-1 alone could achieve in the IGFBP-rich uterine fluid environment.
This reproductive biology connection has implications for understanding both normal fertility and pathological conditions like endometriosis (where ectopic endometrial growth is driven partly by IGF-1 signaling) and endometrial cancer (where aberrant IGF-1R activation is commonly observed). Researchers interested in reproductive peptides may also find connections to Kisspeptin-10 and Gonadorelin, which modulate reproductive function through the hypothalamic-pituitary-gonadal axis.
Wound Healing and Tissue Repair
IGF-1 is a well-established promoter of wound healing, with documented roles in keratinocyte migration, fibroblast proliferation, collagen synthesis, and angiogenesis. Studies using native IGF-1 in wound models have demonstrated a 132% increase in dermal thickness and a 300% increase in connective tissue cells within wound sites.
While direct wound healing studies with des(1-3)IGF-1 are limited, its enhanced potency and tissue-level bioavailability predict superior efficacy in wound environments where IGFBP concentrations are high. Wound fluid contains substantial concentrations of IGFBP-3 and IGFBP-5, and the regulated proteolysis of these binding proteins by wound-associated MMPs is a recognized mechanism for amplifying local IGF-1 signaling during repair. Des(1-3)IGF-1 would bypass this proteolytic requirement, providing immediate maximal IGF-1R activation in the wound bed.
This tissue repair potential connects des(1-3)IGF-1 to a broader family of healing-associated peptides, including BPC-157, TB-500, and GHK-Cu, each of which promotes tissue repair through distinct molecular mechanisms. The BPC-157/TB-500 blend has become particularly popular in the peptide research community for wound and connective tissue applications.
Cartilage and Bone Research
IGF-1 is essential for normal cartilage maintenance and bone formation, acting through IGF-1R on chondrocytes and osteoblasts to stimulate matrix synthesis, cell proliferation, and differentiation. In cartilage research, IGF-1 has been shown to stimulate chondrocyte adhesion to extracellular matrix proteins, promote proliferation of prechondrocytes, and maintain cartilage integrity through enhanced proteoglycan and type II collagen synthesis.
The potential of des(1-3)IGF-1 in these applications lies in its ability to achieve higher effective concentrations at cell surface receptors in the IGFBP-rich environment of cartilage extracellular matrix. Articular cartilage contains significant concentrations of IGFBP-3, -4, and -5, which normally limit IGF-1 access to chondrocyte receptors. By bypassing this regulation, des(1-3)IGF-1 could theoretically enhance the anabolic response in cartilage repair scenarios, though direct comparative studies in cartilage models are still needed.
Research Applications as a Pharmacological Tool
Beyond its potential therapeutic applications, des(1-3)IGF-1 has proven indispensable as a research tool for dissecting IGF signaling biology. Its unique property of retaining full receptor binding while losing IGFBP binding makes it the ideal probe for distinguishing receptor-dependent from binding protein-dependent effects in any experimental system.
This pharmacological approach has been applied in cancer biology to determine whether IGFBP-3's tumor suppressive effects are IGF-dependent or IGF-independent, in bone biology to separate the direct effects of IGF-1R signaling from IGFBP-5-mediated matrix deposition, and in metabolic research to quantify the contribution of IGFBP regulation to tissue-specific IGF-1 sensitivity. Use of the dosing calculator can help researchers plan experiments with appropriate concentrations based on the target tissue and expected IGFBP environment.
Metabolic Effects: Glucose Homeostasis and Insulin Sensitivity
The metabolic effects of des(1-3)IGF-1 deserve separate consideration because they involve a different set of mechanisms from the growth-promoting effects. IGF-1 shares approximately 50% sequence homology with proinsulin, and the IGF-1 receptor shares significant structural similarity with the insulin receptor. This molecular similarity allows IGF-1, particularly when present in high free concentrations, to activate the insulin receptor and produce insulin-like metabolic effects.
In preclinical studies, administration of native IGF-1 reduces blood glucose levels in a dose-dependent manner, with the effect mediated through both IGF-1R-dependent glucose uptake in skeletal muscle and insulin receptor cross-activation. Des(1-3)IGF-1, achieving higher free concentrations per unit dose, produces more pronounced glucose-lowering effects. Guler, Zapf, and Froesch demonstrated in healthy adult volunteers that recombinant native IGF-1 administered intravenously caused significant hypoglycemia, with blood glucose falling by approximately 40% within 30 minutes (Guler HP, et al. New England Journal of Medicine. 1987;317(3):137-140. DOI: 10.1056/NEJM198707163170303).
The glucose-lowering effect of IGF-1 is distinct from that of insulin in several respects. IGF-1 primarily stimulates glucose uptake in skeletal muscle through PI3K/Akt-mediated GLUT4 translocation, similar to insulin. But IGF-1 has a weaker effect on hepatic glucose production suppression compared to insulin, and it does not suppress lipolysis as potently as insulin at equivalent receptor-occupancy levels. The result is a metabolic profile that lowers blood glucose but with a different pattern of tissue effects than equi-glycemic doses of insulin.
For des(1-3)IGF-1 specifically, the rapid onset and short duration of action mean that the glucose-lowering effect occurs as a sharp, transient dip rather than the sustained reduction seen with native IGF-1 or insulin. This temporal profile may actually reduce the overall risk of hypoglycemia-related adverse events compared to longer-acting variants, provided that the peak glucose-lowering effect can be managed - typically by ensuring adequate carbohydrate intake around the time of administration.
Cancer Biology: A Double-Edged Sword
Des(1-3)IGF-1 has played a significant role in cancer research, though the relationship between IGF-1 signaling and cancer is complex and sometimes contradictory. The IGF-1R is expressed on the vast majority of cancer types, and its activation promotes the two hallmarks most relevant to tumor biology: proliferation (through MAPK/ERK) and survival/resistance to apoptosis (through PI3K/Akt). Epidemiological studies have consistently linked higher circulating IGF-1 levels with modestly increased risks of several common cancers, including colorectal (relative risk approximately 1.2-1.4), premenopausal breast (RR ~1.3), and prostate (RR ~1.1-1.4) cancers.
However, the relationship between IGF-1 and cancer is not simply one of promotion. IGFBPs, particularly IGFBP-3, have well-documented tumor-suppressive properties that are at least partly independent of their IGF-1 binding function. IGFBP-3 induces apoptosis in cancer cells through direct interaction with the retinoid X receptor (RXR-alpha) in the nucleus, and through cell-surface interactions that trigger caspase-8-dependent death signaling. These IGF-independent anti-tumor effects of IGFBP-3 mean that the net biological effect of IGF-1 in a tumor depends on the balance between pro-growth IGF-1R signaling and anti-tumor IGFBP-3 signaling.
Des(1-3)IGF-1 is uniquely informative in this context because it activates IGF-1R without sequestering IGFBP-3. When researchers add native IGF-1 to cancer cell cultures, it binds to both IGF-1R (promoting growth) and IGFBP-3 (potentially reducing IGFBP-3's anti-tumor activity). Des(1-3)IGF-1 activates IGF-1R without affecting IGFBP-3 availability, allowing the IGF-1R-dependent and IGFBP-3-dependent effects to be separated experimentally. This approach has been used in breast cancer, prostate cancer, and colorectal cancer cell models to quantify the relative contributions of each pathway.
From a safety perspective, the cancer biology data reinforces caution about prolonged or high-dose use of des(1-3)IGF-1 (or any IGF-1R agonist) in individuals with known or suspected malignancies. The short half-life of des(1-3)IGF-1 limits total receptor exposure time compared to native IGF-1 or LR3, but the higher peak receptor activation could still be relevant in tissues with established neoplastic changes.
Colostrum-Derived IGF-1 DES: Agricultural and Veterinary Research
The original discovery of des(1-3)IGF-1 in bovine colostrum has led to a line of agricultural research exploring whether colostrum-derived growth factors can be used to improve neonatal animal growth and gut development. Studies in neonatal piglets and lambs have examined whether oral administration of colostral extracts enriched in des(1-3)IGF-1 can accelerate gut maturation and improve survival rates in the critical early postnatal period.
Results have been mixed. Oral des(1-3)IGF-1 appears to survive gastric digestion to some degree (the acidic pH of the neonatal stomach actually promotes generation of des(1-3)IGF-1 from native IGF-1 in colostrum), and some studies have shown enhanced intestinal growth with oral supplementation. However, systemically delivered des(1-3)IGF-1 consistently produces larger effects on gut growth than orally delivered material, suggesting that a significant fraction is degraded in the GI tract before absorption. The development of protected delivery systems - enteric coatings, nanoparticle formulations, or mucoadhesive carriers - remains an area of active research in veterinary science.
Skeletal Muscle Research: Satellite Cells and Hypertrophy Signaling
The role of IGF-1 in skeletal muscle biology is well established, and des(1-3)IGF-1 has been used in several key studies to elucidate the mechanisms by which IGF-1 signaling promotes muscle growth. Skeletal muscle expresses IGF-1R on both mature myofibers and satellite cells (the resident stem cells responsible for muscle repair and growth). When IGF-1R is activated on satellite cells, it stimulates their proliferation and subsequent differentiation into new myonuclei that fuse with existing fibers, contributing to hypertrophy through a process called myonuclear addition.
The PI3K/Akt/mTOR pathway downstream of IGF-1R is the central signaling axis for muscle protein synthesis. mTOR activation by Akt leads to phosphorylation of p70S6K and 4E-BP1, two key regulators of ribosomal biogenesis and cap-dependent translation initiation, respectively. This increases the translational capacity and efficiency of the muscle cell, promoting net positive protein balance that manifests as muscle hypertrophy.
Des(1-3)IGF-1 activates this pathway more efficiently than native IGF-1 in cell culture models because more of the peptide reaches the receptor. In L6 myoblasts, des(1-3)IGF-1 at 1 ng/mL produces comparable Akt phosphorylation to native IGF-1 at 10 ng/mL, a 10-fold potency shift consistent with the expected IGFBP evasion advantage. The downstream functional consequences - increased 3H-leucine incorporation (protein synthesis), elevated creatine kinase activity (myogenic differentiation), and enhanced myotube fusion index - all track proportionally with the Akt activation difference.
Whether this cell culture potency advantage translates to proportionally greater muscle growth in vivo is less clear. The in vivo environment is far more complex than a cell culture dish, with multiple cell types, blood flow, innervation, hormonal milieu, and mechanical loading all modulating the response. The lit/lit mouse studies showed enhanced overall growth with des(1-3)IGF-1, but the tissue-specific breakdown suggested that gut and kidney responded more strongly than skeletal muscle, possibly reflecting the higher IGFBP concentrations in the GI tract. Dedicated muscle-specific studies with des(1-3)IGF-1, including measurement of muscle fiber cross-sectional area, satellite cell activation markers, and pathway signaling in intact muscle, are still needed to definitively quantify the in vivo muscle growth advantage. Researchers interested in muscle growth peptide strategies can explore related compounds including AOD-9604 and Fragment 176-191 through the peptide research hub.
Aging and the IGF-1 Axis: Relevance of Des(1-3)IGF-1
The relationship between IGF-1 signaling and aging is one of the most debated topics in gerontology. On one hand, IGF-1 levels decline with age (a phenomenon sometimes called "somatopause"), and this decline is associated with sarcopenia, frailty, cognitive decline, and impaired tissue repair. On the other hand, reduced IGF-1 signaling is one of the most conserved longevity-promoting pathways across species, from nematodes (daf-2 mutants) to flies (chico mutants) to mice (GH receptor knockout mice). This apparent paradox - that lower IGF-1 may promote both aging-related dysfunction and longevity - remains unresolved.
Des(1-3)IGF-1 offers an interesting lens through which to view this paradox. If the goal is to restore tissue-specific IGF-1 signaling in an aging organism without chronically elevating systemic IGF-1 levels, a short-acting, locally administered IGF-1 variant could theoretically provide the tissue repair benefits of IGF-1R activation while minimizing the sustained systemic exposure that may accelerate aging and age-related disease. This pulsatile approach mirrors the natural pattern of GH secretion, which occurs in discrete pulses rather than continuous release, and may explain why pulsatile GH secretion promotes growth while continuous GH exposure promotes pathology.
Researchers exploring the intersection of growth factor signaling and aging may find des(1-3)IGF-1 relevant alongside other longevity-associated peptides and compounds, including Epithalon (telomerase activation), FOXO4-DRI (senolytic targeting), SS-31 (mitochondrial protection), Humanin (mitochondrial-derived peptide), and MOTS-c (mitochondrial ORF of the twelve S rRNA). Each of these compounds interacts with different aspects of the aging process, and their combined study is advancing our understanding of how peptide-based interventions might modulate aging trajectories. The biohacking hub provides additional context on anti-aging peptide research.
Comparison: DES vs LR3 vs Native
Three forms of IGF-1 dominate the research literature and the peptide market: native IGF-1 (mecasermin), IGF-1 LR3 (Long R3 IGF-1), and IGF-1 DES (Des(1-3)IGF-1). Each represents a different approach to modifying IGF-1 pharmacology, and understanding their distinct profiles is essential for selecting the appropriate variant for a given research question or clinical application.
Structural Differences at a Glance
The three variants differ in their N-terminal modifications, which determine their IGFBP binding characteristics and pharmacokinetic profiles:
Native IGF-1 (mecasermin): The full 70-amino-acid human IGF-1 sequence. This is the physiological form, produced primarily by the liver under growth hormone stimulation. It has full affinity for all six IGFBPs and the IGF-1 receptor, with a circulating half-life of 12-16 hours when bound to the ternary complex.
IGF-1 LR3 (Long R3 IGF-1): An 83-amino-acid synthetic analogue with two modifications: an arginine substituted for glutamic acid at position 3, and a 13-amino-acid extension peptide added to the N-terminus. These changes reduce IGFBP binding by approximately 100-fold (similar in magnitude to des(1-3)IGF-1) while extending the peptide's half-life to approximately 20-30 hours due to altered clearance kinetics and resistance to proteolytic degradation.
IGF-1 DES (Des(1-3)IGF-1): A 67-amino-acid truncated form lacking the first three N-terminal residues (Gly-Pro-Glu). IGFBP binding is reduced by approximately 99%, and the half-life is shortened to 20-30 minutes due to rapid renal clearance in the absence of binding protein protection.
Comprehensive Comparison Table
| Property | Native IGF-1 | IGF-1 LR3 | IGF-1 DES |
|---|---|---|---|
| Amino Acids | 70 | 83 | 67 |
| Molecular Weight | ~7,649 Da | ~9,111 Da | ~7,372 Da |
| Modification | None (wild-type) | Glu3Arg + 13-aa N-terminal extension | N-terminal truncation (del 1-3) |
| IGF-1R Binding Affinity | Full (Kd ~1 nM) | ~2-3x native | Full (equivalent to native) |
| IGFBP Binding | Full (high affinity) | ~1% of native | ~1% of native |
| Biological Potency (in vitro) | 1x (reference) | ~2-3x | ~10x |
| Half-Life | 12-16 hours (bound) | 20-30 hours | 20-30 minutes |
| Occurs Naturally | Yes (liver, tissues) | No (synthetic only) | Yes (colostrum, brain, uterus) |
| FDA-Approved Form | Yes (mecasermin/Increlex) | No | No |
| Hypoglycemia Risk | Moderate (dose-dependent) | Moderate-High | High (rapid onset) |
| Primary Duration of Action | Sustained (hours) | Prolonged (day+) | Rapid, brief (minutes) |
Potency Differences Explained
At first glance, the potency rankings seem counterintuitive. IGF-1 LR3 and IGF-1 DES both show similar reductions in IGFBP binding (about 99%), yet DES is roughly 3-5 times more potent than LR3 in most cell-based assays. Why?
The answer lies in receptor pharmacology. While both variants escape IGFBP sequestration, IGF-1 LR3's additional 13-amino-acid extension creates a slightly larger molecule that may experience minor steric constraints at the receptor binding site. More significantly, the LR3 extension appears to partially offset the IGFBP evasion by providing alternative serum protein binding interactions that reduce the free fraction somewhat. The net result is that LR3 achieves about 2-3x the potency of native IGF-1, while DES achieves the full theoretical benefit of IGFBP escape at approximately 10x.
But potency isn't everything. The dramatically different half-lives of these variants mean they occupy fundamentally different pharmacological niches.
Pharmacokinetic Profiles: When Duration Matters
IGF-1 LR3: The Marathon Runner
With a half-life of 20-30 hours, IGF-1 LR3 provides sustained IGF-1R activation over an extended period. A single administration can maintain elevated IGF signaling for 24-48 hours, making it suitable for applications where continuous receptor stimulation is desired. In cell culture, LR3 is commonly added to media at the time of plating and provides growth factor support throughout the culture period without the need for repeated additions. The extended half-life is attributed to resistance to proteolytic degradation conferred by the N-terminal extension and the substitution at position 3.
IGF-1 DES: The Sprinter
Des(1-3)IGF-1's 20-30 minute half-life creates a pharmacokinetic profile that favors pulsatile, localized effects. After administration, peak receptor activation occurs rapidly, and the peptide is cleared within 1-2 hours. This makes it particularly interesting for applications requiring precisely timed IGF-1R activation without sustained systemic exposure - a profile analogous to the pulsatile secretion patterns of growth hormone itself.
The short half-life also creates a practical advantage for in vivo research: the ability to administer des(1-3)IGF-1 at specific time points and then analyze tissues at known intervals after peak exposure, without the complication of continued receptor activation from residual peptide.
Native IGF-1: The Regulated Middle Ground
Mecasermin (recombinant human IGF-1) occupies the middle ground. Its 12-16 hour half-life in the ternary complex provides sustained but not indefinite signaling, and its full IGFBP binding means that the body's normal regulatory machinery governs its bioavailability. This makes it the safest of the three from a regulatory standpoint, which is reflected in its status as the only FDA-approved IGF-1 product (marketed as Increlex for the treatment of severe primary IGF-1 deficiency).
Application-Specific Recommendations
Research Application Guide
- Cell culture maintenance: IGF-1 LR3 is preferred for its stability in media and extended bioactivity
- Acute signaling studies: IGF-1 DES provides a clean, time-limited pulse of receptor activation
- IGFBP biology research: Use both DES and native IGF-1 in parallel to separate receptor-dependent from IGFBP-dependent effects
- Animal growth studies: Native IGF-1 for physiological modeling; LR3 for sustained anabolic effects; DES for tissue-specific targeting
- Gut/mucosal applications: DES has the strongest preclinical data for intestinal tissue growth
Insulin Receptor Cross-Reactivity Comparison
All three IGF-1 variants retain some cross-reactivity with the insulin receptor, but the clinical significance differs markedly. Native IGF-1, with 95-99% bound to IGFBPs, rarely achieves free concentrations sufficient for meaningful insulin receptor activation under normal physiological conditions. IGF-1 LR3 and DES, both present in predominantly free form, can activate insulin receptors at lower total doses.
The practical difference is timing. IGF-1 LR3's sustained presence means that insulin receptor cross-activation, if it occurs, persists for hours. IGF-1 DES's rapid clearance means that any insulin-like effects are transient, typically resolving within 30-60 minutes. This has implications for hypoglycemia risk management, which differs substantially between the two variants.
The "Best" Variant Depends on the Question
There is no universally superior IGF-1 variant. The choice depends entirely on the research question or therapeutic goal:
If you need sustained, systemic IGF-1R activation with minimal dosing frequency, IGF-1 LR3 is the appropriate choice. Its long half-life provides all-day receptor engagement from a single dose, and its resistance to proteolytic degradation makes it stable in most experimental and clinical contexts.
If you need rapid, high-potency, localized IGF-1R activation with minimal systemic persistence, IGF-1 DES is the better option. Its 10-fold potency advantage and short half-life create a pharmacological profile that favors tissue-targeted effects with a predictable offset.
If you need physiological modeling of IGF-1 signaling with full regulatory context, native IGF-1 (mecasermin) provides the most faithful representation of how IGF-1 behaves in the human body, complete with IGFBP regulation, ternary complex formation, and normal clearance kinetics.
Many researchers maintain all three variants in their toolkit, selecting the one that best matches each experimental design. For those building a broader peptide research program, understanding these IGF-1 variants alongside related growth-promoting peptides like CJC-1295 DAC, tesamorelin, and IGF-1 LR3 provides a comprehensive foundation in growth factor pharmacology. The drug comparison hub offers additional context on how these compounds relate to each other.
Tissue-Specific Considerations in Variant Selection
The choice of IGF-1 variant also depends heavily on the target tissue and its local IGFBP environment. Different tissues express different combinations and concentrations of IGFBPs, which means the effective potency advantage of des(1-3)IGF-1 and LR3 varies by anatomical location.
Gastrointestinal Tract
The GI mucosa expresses high levels of IGFBP-3 and IGFBP-5, creating an environment where native IGF-1 is heavily buffered. This makes the gut one of the tissues where des(1-3)IGF-1 shows the most pronounced advantage over native IGF-1, consistent with the dramatic gut growth seen in the Lemmey et al. resection studies. The intestinal epithelium turns over every 3-5 days, and IGF-1 signaling through the crypt stem cell compartment is a major driver of this renewal. Des(1-3)IGF-1's ability to bypass IGFBP regulation in this context may produce disproportionately strong effects on crypt cell proliferation and mucosal regeneration.
Skeletal Muscle
Skeletal muscle expresses IGFBP-4 and IGFBP-5 in the extracellular matrix surrounding muscle fibers, along with the IGFBP-4 protease PAPP-A. In this tissue, IGFBPs play a nuanced role: IGFBP-5 bound to the extracellular matrix can actually potentiate IGF-1 action by concentrating the growth factor near its receptor. Des(1-3)IGF-1 cannot benefit from this IGFBP-5-mediated potentiation, which may partially offset its free-fraction advantage in muscle tissue specifically. LR3, which retains very low but non-zero IGFBP binding, may interact differently with the muscle IGFBP environment, though this has not been directly tested.
Bone and Cartilage
The bone microenvironment is particularly rich in IGFBPs, with IGFBP-3, -4, and -5 all present in significant concentrations in the bone matrix. IGF-1 stored in the bone matrix is released during bone resorption and acts locally on osteoblasts to couple bone remodeling. Des(1-3)IGF-1 would bypass this matrix-associated IGFBP regulation entirely, potentially producing rapid osteoblast stimulation without the normal coupling to resorption signals. Whether this represents an advantage or a disruption of normal bone remodeling homeostasis depends on the clinical context.
Central Nervous System
The brain represents a unique case because des(1-3)IGF-1 occurs naturally there. IGFBP-2 is the dominant binding protein in cerebrospinal fluid, and its concentrations are considerably lower than those of IGFBP-3 in serum. The brain may already operate in a relatively low-IGFBP environment for IGF-1 signaling, which would narrow the potency gap between des(1-3)IGF-1 and native IGF-1 in neural tissue. However, the endogenous production of des(1-3)IGF-1 in the brain suggests that even the modest IGFBP buffering present in the CNS is significant enough that the body selectively produces the IGFBP-resistant form for local use. Researchers interested in neuroprotective peptide strategies may find value in exploring IGF-1 DES alongside dedicated neuropeptides like Semax and Pinealon.
Practical Dosing Considerations Across Variants
The three IGF-1 variants require fundamentally different dosing strategies due to their distinct pharmacokinetic profiles. This section provides general guidance based on published preclinical protocols and should not be interpreted as clinical dosing recommendations, as no human dose-finding studies have been conducted for des(1-3)IGF-1 or LR3.
For native IGF-1 (mecasermin), the FDA-approved dosing in children with severe primary IGF-1 deficiency is 0.04-0.12 mg/kg given twice daily by subcutaneous injection. This dosing achieves physiological to slightly supraphysiological IGF-1 levels, with the 12-16 hour half-life allowing twice-daily dosing to maintain relatively steady-state concentrations. The extensive clinical trial data for mecasermin provides the most strong safety information of any IGF-1 variant.
IGF-1 LR3, in research applications, has typically been used at doses ranging from 20-100 mcg/day in animal studies, scaled by body weight. Its 20-30 hour half-life means that once-daily dosing achieves near-steady-state conditions within 2-3 days. Researchers commonly use LR3 in cell culture at concentrations of 10-100 ng/mL in serum-supplemented media.
Des(1-3)IGF-1, with its 20-30 minute half-life, requires a different conceptual approach. Rather than aiming for sustained receptor occupancy, dosing is typically designed around acute activation windows. In animal research protocols, des(1-3)IGF-1 has been administered by continuous subcutaneous infusion (to maintain steady-state levels) or by multiple daily bolus injections. In cell culture, it is typically added at 1-10 ng/mL - roughly 10-fold lower concentrations than native IGF-1 for equivalent effects.
The dosing calculator on FormBlends provides additional guidance for researchers planning experiments with these compounds, incorporating published dose-response data from the preclinical literature.
Stability and Handling Across Variants
Practical laboratory considerations also differ among the three variants. Native IGF-1 is relatively stable when stored lyophilized at -20C but degrades within days at room temperature in solution. IGF-1 LR3 benefits from enhanced stability due to its N-terminal extension, which provides some protection against N-terminal aminopeptidase degradation. Des(1-3)IGF-1, lacking three N-terminal residues, is not subject to N-terminal degradation pathways but may be slightly less stable than native IGF-1 overall due to its smaller size and reduced ability to form stabilizing complexes with serum proteins.
All three variants should be reconstituted in acidified water or dilute acetic acid (0.1M acetic acid is commonly used) to prevent aggregation and maintain bioactivity. Repeated freeze-thaw cycles should be avoided. Working aliquots stored at 4C should be used within 1-2 weeks. These handling requirements are standard for small peptides and growth factors and apply equally to other compounds in the research toolkit, from BPC-157 to Epithalon.
The Insulin/IGF-1 Receptor Hybrid Question
A frequently overlooked complication in comparing IGF-1 variants is the existence of insulin/IGF-1 hybrid receptors. In many tissues, particularly skeletal muscle and adipose tissue, hybrid receptors composed of one insulin receptor hemireceptor and one IGF-1R hemireceptor are expressed at significant levels. These hybrid receptors have binding properties that differ from either homodimeric receptor: they bind IGF-1 with high affinity (similar to IGF-1R homodimers) but bind insulin with reduced affinity compared to insulin receptor homodimers.
The significance of hybrid receptors for des(1-3)IGF-1 pharmacology is largely unexplored. Because des(1-3)IGF-1 binds IGF-1R with full affinity, it presumably also binds hybrid receptors with high affinity, as the IGF-1-binding surface on the hybrid receptor is contributed by the IGF-1R hemireceptor. However, the signaling output of hybrid receptor activation may differ from that of IGF-1R homodimer activation, as the intracellular domains of the insulin receptor and IGF-1R hemireceptors can recruit different adaptor proteins and activate different signaling pathways.
In tissues where hybrid receptors predominate, the biological response to des(1-3)IGF-1 may be a blend of "pure" IGF-1R signaling and "hybrid" signaling with characteristics of both IGF-1 and insulin pathways. This could contribute to the metabolic effects (glucose uptake, lipid metabolism) seen with high-dose IGF-1 administration and may partially explain tissue-specific variations in the response to des(1-3)IGF-1 versus native IGF-1.
The IGF-2 Connection
Any complete discussion of IGF-1 variant pharmacology must acknowledge the role of IGF-2, the second member of the IGF ligand family. IGF-2 binds to IGF-1R with approximately 2-10 fold lower affinity than IGF-1, but it is present in much higher concentrations in many tissues, particularly during embryonic development. IGF-2 also binds to the IGF-2R (also known as the mannose-6-phosphate receptor), which functions primarily as a clearance receptor rather than a signaling receptor.
The relevance of IGF-2 to des(1-3)IGF-1 research is twofold. First, IGF-2 binds to some of the same IGFBPs as IGF-1 (particularly IGFBP-2, -3, and -6), creating competition for binding protein binding that can influence free IGF-1 levels in tissues where both ligands are present. Des(1-3)IGF-1 is immune to this competitive dynamic because it doesn't bind IGFBPs, but the competition between native IGF-1 and IGF-2 for IGFBP binding is an important variable in experimental systems where both ligands are present. Second, IGF-2 can activate IGF-1R, and its contribution to total IGF-1R signaling in any tissue must be considered when interpreting the effects of exogenous des(1-3)IGF-1 administration.
Researchers studying IGF-1 DES should be aware that standard IGF-1 immunoassays do not cross-react with IGF-2, and that total IGF-1R signaling in a tissue reflects contributions from both endogenous IGF-1, endogenous IGF-2, and any exogenous des(1-3)IGF-1. Pathway-level readouts (phospho-Akt, phospho-ERK) capture the sum of all these inputs, while ligand-level measurements require specific assays for each molecule.
Economic and Accessibility Considerations
The practical availability of IGF-1 variants differs substantially, which affects research accessibility and cost.
Native IGF-1 (mecasermin) is available as an FDA-approved pharmaceutical product (Increlex), but its cost is extremely high - typically $5,000-$10,000+ per month at therapeutic doses for IGF-1 deficiency in children. Research-grade recombinant human IGF-1 is available from multiple suppliers at more accessible prices for laboratory use.
IGF-1 LR3 is the most widely available of the modified variants, with numerous research peptide suppliers offering it in both lyophilized and solution formats. Its stability and long half-life make it popular for cell culture applications, where it has become a standard media supplement for many IGF-1-dependent cell lines. Research-grade LR3 is available from specialty suppliers including those focused on cell culture reagents.
Des(1-3)IGF-1 is less widely available than LR3 but can be obtained from several research peptide suppliers and specialty growth factor companies. Its shorter half-life and more specialized applications mean lower overall demand compared to LR3, which is reflected in fewer commercial sources. Quality and purity vary between suppliers, and researchers should verify the identity and activity of commercial des(1-3)IGF-1 preparations using appropriate analytical methods (mass spectrometry, cell-based bioassays) before use in experiments.
For researchers evaluating these compounds, FormBlends' IGF-1 DES and IGF-1 LR3 product pages provide additional information on available formulations and specifications.
Safety Considerations

Des(1-3)IGF-1 has no completed human clinical trials, no FDA approval, and no established safety profile in humans. All safety assessments are extrapolated from preclinical animal data, the known pharmacology of the IGF-1 signaling pathway, and the clinical safety record of related compounds like mecasermin (Increlex). This section examines the identified and theoretical risks based on available evidence.
Hypoglycemia: The Primary Acute Risk
The most significant acute safety concern with des(1-3)IGF-1 is hypoglycemia. IGF-1 shares structural homology with insulin and activates the insulin receptor at high concentrations. Because des(1-3)IGF-1 achieves higher free concentrations than equivalent doses of native IGF-1, the threshold for insulin receptor cross-activation is lower, and the onset of any hypoglycemic effect is faster.
Data from mecasermin (Increlex) provides the closest clinical reference point. In the European IGFD Registry, hypoglycemia was reported in approximately 49% of patients treated with recombinant IGF-1, with severe hypoglycemia (requiring assistance) occurring in about 5.7% of cases. These figures apply to native IGF-1, which has full IGFBP buffering. Des(1-3)IGF-1, lacking this buffering, would be expected to produce more rapid-onset and potentially more severe hypoglycemic episodes at equivalent molar doses.
The short half-life of des(1-3)IGF-1 is actually a partial mitigating factor here. While hypoglycemia onset may be faster, the duration of the hypoglycemic episode should be shorter than with native IGF-1 or LR3, resolving within 30-60 minutes as the peptide is cleared. This contrasts with IGF-1 LR3, where hypoglycemia from insulin receptor cross-activation could theoretically persist for hours.
Hypoglycemia Risk Factors
- Concurrent insulin use dramatically increases the risk of severe, potentially life-threatening hypoglycemia
- Fasting or carbohydrate restriction at the time of administration amplifies the hypoglycemic effect
- Higher doses produce more pronounced insulin receptor cross-activation
- The rapid onset (within minutes) means that traditional hypoglycemia countermeasures may need to be immediately available
- Individual variation in insulin sensitivity, IGFBP levels, and receptor expression creates unpredictable dose-response relationships
Growth Factor-Related Oncological Concerns
Any molecule that activates the IGF-1 receptor raises theoretical concerns about cancer promotion. The IGF-1R/PI3K/Akt signaling axis is one of the most frequently activated pathways in human cancers, and epidemiological data consistently link elevated circulating IGF-1 levels with modestly increased risks of colorectal, breast, and prostate cancer. The association is not with cancer initiation per se, but with promotion and progression of existing neoplastic cells.
Des(1-3)IGF-1's enhanced potency and escape from IGFBP regulation theoretically amplify this concern. IGFBPs, particularly IGFBP-3, have well-documented IGF-independent anti-proliferative and pro-apoptotic effects on cancer cells. By bypassing IGFBP sequestration, des(1-3)IGF-1 simultaneously increases IGF-1R-mediated mitogenic signaling and removes the IGFBP-mediated tumor-suppressive counterbalance.
It is essential to note, however, that no direct evidence exists linking des(1-3)IGF-1 administration with cancer development in any animal model. The concern is theoretical, based on pathway biology rather than observed outcomes. The short half-life of des(1-3)IGF-1 also limits the total duration of receptor exposure compared to sustained IGF-1 elevation, which may reduce the relevance of this concern in practice.
Localized Tissue Overgrowth
The potent mitogenic effects of des(1-3)IGF-1 raise concerns about localized tissue overgrowth with repeated administration, particularly at injection sites. IGF-1 is a known driver of organ growth - the acromegalic phenotype seen with growth hormone excess is mediated in part through elevated IGF-1 signaling. While des(1-3)IGF-1's short half-life limits systemic exposure, concentrated local delivery could theoretically produce disproportionate tissue growth at the site of administration.
This concern is particularly relevant in the context of subcutaneous or intramuscular injection, where high local concentrations may persist in the tissue depot longer than the plasma half-life suggests. The formation of localized fibrotic nodules or tissue hypertrophy at injection sites has been reported anecdotally, though controlled studies documenting this effect are lacking.
Cardiovascular Considerations
IGF-1 signaling affects cardiovascular function through multiple mechanisms. IGF-1R is expressed on cardiomyocytes, vascular smooth muscle cells, and endothelial cells, where it promotes cell survival, angiogenesis, and cardiac remodeling. Both deficiency and excess of IGF-1 signaling have been associated with cardiovascular pathology. Chronic IGF-1 excess can promote cardiac hypertrophy, and in acromegaly, cardiomyopathy is a recognized complication.
The brief exposure profile of des(1-3)IGF-1 makes chronic cardiovascular effects unlikely with intermittent use. However, the acute hemodynamic effects of rapidly elevated free IGF-1 - including potential vasodilation, changes in cardiac contractility, and acute blood pressure fluctuations - remain uncharacterized in humans.
Absence of Pharmacokinetic Data in Humans
No formal pharmacokinetic studies of des(1-3)IGF-1 have been conducted in humans. All half-life estimates, volume of distribution projections, and clearance predictions are extrapolated from animal models and from the behavior of structurally related compounds. This represents a significant knowledge gap, as species differences in IGFBP concentrations, renal clearance rates, and tissue receptor density could produce pharmacokinetic behavior in humans that differs substantially from animal predictions.
The absence of human PK data also means that dosing guidance cannot be established with any clinical rigor. Doses used in animal research are scaled by body weight but have not been validated for safety or efficacy in humans. For those seeking structured approaches to peptide dosing, the FormBlends dosing calculator provides general guidance based on available preclinical literature, though it cannot substitute for proper clinical assessment.
Drug Interactions
The most clinically significant potential interaction involves concurrent insulin or insulin secretagogue use. Both IGF-1 and insulin lower blood glucose through overlapping but distinct mechanisms, and their combined effect on blood glucose is additive or potentially complementary. Use of des(1-3)IGF-1 alongside exogenous insulin carries a substantial risk of severe, potentially life-threatening hypoglycemia that exceeds the risk of either agent alone.
Other potential interactions are less well characterized but include:
- Growth hormone: GH stimulates hepatic IGF-1 production and IGFBP secretion. Co-administration with des(1-3)IGF-1 could produce complex, unpredictable effects on the GH-IGF axis feedback loops.
- Glucocorticoids: These suppress IGF-1 signaling and increase IGFBP-1 levels. Des(1-3)IGF-1 would be unaffected by the IGFBP-1 increase but would still face glucocorticoid-mediated resistance at the receptor level.
- GLP-1 receptor agonists: Compounds like semaglutide and tirzepatide affect glucose homeostasis through incretin pathways that interact with the insulin-IGF axis. The combined metabolic effects have not been studied.
Regulatory Status
Des(1-3)IGF-1 is not approved by any regulatory agency for human therapeutic use. It is classified as a research peptide and is legally available for in vitro and animal research purposes. It is included on the World Anti-Doping Agency (WADA) prohibited list under the category of peptide hormones, growth factors, and related substances. Athletes subject to anti-doping testing should be aware that des(1-3)IGF-1 is a prohibited substance both in and out of competition.
Acromegaly-Like Effects: Lessons from Chronic GH/IGF-1 Excess
Acromegaly, caused by chronic GH excess from pituitary tumors, provides a clinical model for what sustained IGF-1R overactivation looks like in humans. The features include soft tissue swelling, enlargement of the hands, feet, and jaw, organ hypertrophy (particularly cardiac), insulin resistance progressing to diabetes, and increased risk of colon polyps and colorectal cancer. These manifestations develop over years to decades of chronic exposure to elevated IGF-1 levels.
The relevance of acromegaly to des(1-3)IGF-1 safety is indirect but instructive. Des(1-3)IGF-1's short half-life means that sustained, acromegaly-like exposure is unlikely with intermittent dosing. However, repeated daily administration at high doses could theoretically produce cumulative tissue effects, particularly in rapidly dividing tissues like the intestinal mucosa. The gut-selective growth seen in preclinical studies with des(1-3)IGF-1 is actually reminiscent of the colonic mucosal proliferation seen in acromegaly, suggesting that this tissue may be particularly vulnerable to IGF-1R-driven growth.
The cardiac effects of chronic IGF-1 excess are also worth considering. Acromegalic cardiomyopathy involves both concentric hypertrophy and eventual systolic/diastolic dysfunction. While des(1-3)IGF-1's brief exposure profile makes this unlikely with typical research protocols, it represents a theoretical concern with chronic high-dose regimens that should be monitored in any long-term animal study.
Injection Site Reactions and Local Effects
As with all peptide injectables, local reactions at the injection site are possible with des(1-3)IGF-1. These may include redness, swelling, pain, and itching. More concerning is the possibility of localized tissue overgrowth at sites of repeated injection, given the peptide's potent mitogenic properties. While formal studies documenting this effect with des(1-3)IGF-1 specifically are lacking, analogous effects have been reported with other locally administered growth factors, and the theoretical basis for concern is strong.
The short half-life of des(1-3)IGF-1 creates an interesting local pharmacokinetic scenario. After subcutaneous injection, the peptide concentrations at the injection site are initially very high (potentially 100-1,000 times the systemic concentration) and decline rapidly as the peptide diffuses into the surrounding tissue and vasculature. This creates a steep concentration gradient that exposes cells immediately adjacent to the injection site to growth factor levels far above those experienced by distant tissues. Repeated injection at the same site could produce cumulative local effects that are not representative of systemic exposure.
Rotating injection sites, using dilute concentrations, and avoiding repeated administration to the same tissue location are standard precautions that may reduce the risk of localized overgrowth. These are the same injection site management principles that apply to insulin administration and other peptide therapies.
Immune Considerations
Des(1-3)IGF-1 is a modified form of an endogenous human protein, which reduces but does not eliminate the potential for immunogenic responses. The removal of three amino acids creates a novel N-terminal sequence that could, in theory, be recognized as non-self by the adaptive immune system. However, the small size of the modification and the structural similarity to the native form make clinically significant immunogenicity unlikely.
With recombinant native IGF-1 (mecasermin), anti-IGF-1 antibodies have been detected in a small percentage of treated patients (roughly 5-10%), though these antibodies have generally not been neutralizing and have not been associated with adverse clinical effects. Whether des(1-3)IGF-1 would produce a similar or different antibody response is unknown, as no human immunogenicity studies have been conducted.
Monitoring and Risk Mitigation Strategies
For researchers conducting animal studies with des(1-3)IGF-1, the following monitoring parameters are recommended based on the known pharmacology and safety profile of IGF-1 signaling:
- Blood glucose: Monitor before and at 15, 30, and 60 minutes after each dose to characterize the hypoglycemic response
- Body weight and organ weights: Track regularly to detect disproportionate organ growth, particularly of the gut, kidneys, and spleen
- Histopathology: Examine tissues for hyperplasia, dysplasia, or neoplastic changes at study termination
- Serum IGF-1 and IGFBP levels: Monitor to assess potential feedback effects on the endogenous IGF axis
- Insulin levels and glucose tolerance: Evaluate for changes in insulin sensitivity that may persist beyond the acute dosing window
- Injection site examination: Inspect for fibrosis, hypertrophy, or other local tissue changes
These monitoring parameters should be adapted to the specific study design, duration, and endpoints. For longer-term studies, periodic assessment of cardiac function (echocardiography in larger animal models) and cancer screening (where applicable) may also be warranted.
Critical Safety Summary
Des(1-3)IGF-1 has no established safety profile in humans. The primary acute risk is hypoglycemia, which can be severe and rapid in onset. Theoretical long-term concerns include growth factor-mediated cancer promotion and localized tissue overgrowth. The absence of human pharmacokinetic, pharmacodynamic, and safety data means that any use beyond controlled research settings carries unknown and unquantifiable risks. Always consult a qualified healthcare provider before considering any peptide therapy. The free assessment can help connect you with appropriate clinical guidance.
Comparison of Safety Profiles Across IGF-1 Variants
| Safety Parameter | Native IGF-1 (Mecasermin) | IGF-1 LR3 | IGF-1 DES |
|---|---|---|---|
| Human Safety Data | Extensive (FDA-approved) | None (research only) | None (research only) |
| Hypoglycemia Risk | Moderate, dose-dependent | Moderate-High, prolonged | High, rapid but brief |
| Duration of Hypoglycemia | Hours (matches half-life) | Potentially 12+ hours | 30-60 minutes |
| Cancer Risk (theoretical) | Low-Moderate (epidemiological data available) | Unknown (sustained exposure) | Unknown (high peak, short duration) |
| Immunogenicity | Low (5-10% develop non-neutralizing antibodies) | Unknown | Unknown |
| Injection Site Effects | Mild pain, occasional lipohypertrophy | Unknown | Theoretical localized overgrowth risk |
| Organ Hypertrophy Risk | Low at approved doses | Unknown (sustained signaling) | Gut-selective in animal studies |
| Drug Interactions | Well-characterized | Poorly characterized | Poorly characterized |
This safety comparison highlights a recurring theme: native IGF-1 is the only variant with a characterized human safety profile. For des(1-3)IGF-1 and IGF-1 LR3, safety assessments are necessarily based on extrapolation from preclinical data and the known pharmacology of the IGF-1 signaling pathway. Researchers and clinicians should approach these variants with appropriate caution and, where possible, design studies that include the safety monitoring parameters outlined above.
Lessons from Mecasermin (Increlex) Clinical Experience
The clinical safety record of mecasermin (recombinant human IGF-1, marketed as Increlex) provides the most relevant human safety data for extrapolating potential des(1-3)IGF-1 risks. Increlex has been approved since 2005 for the treatment of severe primary IGF-1 deficiency (formerly called Laron syndrome) in children, and its post-marketing surveillance database now spans nearly two decades of clinical experience.
The most commonly reported adverse events with mecasermin include hypoglycemia (49% of patients in the European IGFD Registry, with 5.7% experiencing severe episodes), injection site reactions (16-20%), headache (11-15%), tonsillar and adenoidal hypertrophy (7-10%), and lipohypertrophy at injection sites (5-8%). Less common but clinically significant events include intracranial hypertension (pseudotumor cerebri), scoliosis progression, and allergic reactions.
The tonsillar and adenoidal hypertrophy finding is particularly interesting because it reflects IGF-1R-driven tissue growth in the oropharyngeal lymphoid tissue, consistent with the gut-selective growth effects seen with des(1-3)IGF-1 in animal studies. Both the gut mucosa and the oropharyngeal lymphoid tissue are IGF-1-responsive tissues with high turnover rates, and their disproportionate growth response to systemic IGF-1 exposure suggests that these tissues are particularly sensitive to IGF-1R activation.
For des(1-3)IGF-1, the mecasermin safety data suggests that hypoglycemia would be the most likely acute adverse event, with the onset potentially faster and the severity potentially greater per unit dose due to the higher free fraction. The tissue-specific growth effects (tonsillar hypertrophy, gut mucosal hyperplasia) would be expected to be more pronounced with des(1-3)IGF-1 at equivalent total doses, given its enhanced tissue-level bioavailability. However, the shorter half-life might partially offset this by reducing the total cumulative receptor exposure over a 24-hour period.
Contraindications and Precautions: Extrapolated from IGF-1 Pharmacology
Based on the known pharmacology of IGF-1 signaling and the clinical safety experience with mecasermin, the following conditions would likely represent contraindications or require special precautions for des(1-3)IGF-1 use in any future clinical context:
- Active malignancy or history of cancer: IGF-1R activation promotes proliferation and survival of cancer cells. Patients with active malignancies or a recent cancer history should not receive IGF-1R agonists, including des(1-3)IGF-1.
- Closed epiphyses (adults on growth-promoting regimens): While des(1-3)IGF-1 can stimulate bone growth, in adults with closed growth plates, the growth-promoting effects would be limited to soft tissues and could contribute to acromegalic-type changes.
- Concurrent insulin therapy: The additive hypoglycemic risk of combined insulin and des(1-3)IGF-1 would require extremely careful blood glucose monitoring and dose adjustment. This combination should be avoided outside of controlled clinical research settings.
- Diabetic retinopathy: Active proliferative diabetic retinopathy involves VEGF-driven neovascularization, and IGF-1 signaling has been implicated as a cofactor in retinal neovascularization. Des(1-3)IGF-1 could theoretically worsen proliferative retinopathy, though this has not been directly studied.
- Pregnancy: IGF-1 signaling plays critical roles in embryonic and fetal development, and exogenous IGF-1R activation during pregnancy could disrupt normal growth factor balance. The uterine presence of endogenous des(1-3)IGF-1 suggests that the reproductive tract is particularly sensitive to this signaling pathway.
- Renal impairment: Altered renal clearance would extend the half-life and potentially the hypoglycemic window of des(1-3)IGF-1, requiring dose adjustment in patients with reduced kidney function.
These contraindications are theoretical extrapolations from IGF-1 pathway biology and the mecasermin safety database. No clinical data specific to des(1-3)IGF-1 exists to confirm or refute any of them. Healthcare providers considering any peptide intervention should conduct thorough individual risk assessments. The free assessment tool provides an initial framework for such evaluations.
Ethical and Regulatory Framework
The use of des(1-3)IGF-1 outside of controlled research settings raises ethical and regulatory considerations that researchers and clinicians should be aware of. As an unapproved substance with no established human safety profile, des(1-3)IGF-1 does not meet the evidentiary standards required for off-label prescribing, informed consent, or clinical research without appropriate regulatory oversight.
In the United States, the use of des(1-3)IGF-1 in human research would require an Investigational New Drug (IND) application to the FDA, Institutional Review Board (IRB) approval of the research protocol, and appropriate informed consent from research participants. These regulatory requirements exist to protect research subjects from the risks inherent in testing substances without established safety profiles.
The WADA prohibition of des(1-3)IGF-1 in competitive sports applies to all athletes subject to anti-doping testing. Detection methods for IGF-1 analogues have advanced considerably in recent years, with mass spectrometry-based assays capable of distinguishing between endogenous des(1-3)IGF-1 (present in trace amounts in brain tissue) and exogenously administered material based on pharmacokinetic profiles and isotope ratios.
Future Research Directions and Unresolved Questions
Despite three decades of preclinical research, several fundamental questions about des(1-3)IGF-1 remain unanswered. Addressing these gaps would significantly advance the field and inform any potential clinical development.
Human Pharmacokinetics
The most critical data gap is the absence of human PK studies. The estimated 20-30 minute half-life is extrapolated from animal models, but species differences in renal clearance, tissue distribution volumes, and residual IGFBP binding could produce significantly different pharmacokinetics in humans. A single-dose PK study in healthy volunteers, measuring plasma des(1-3)IGF-1 concentrations at frequent intervals after subcutaneous injection, would provide the foundational data needed to design any subsequent efficacy study. Such a study would also characterize the glucose-lowering time course and provide the first human safety data under controlled conditions.
Tissue Distribution Studies
The tissue distribution of des(1-3)IGF-1 after systemic administration has not been comprehensively mapped, even in animals. Radiolabeled or fluorescently tagged des(1-3)IGF-1 studies would reveal which tissues preferentially accumulate the peptide, whether it crosses the blood-brain barrier efficiently, and how its distribution profile compares to native IGF-1 and LR3. These data would inform tissue-targeting strategies and help predict which therapeutic applications are most feasible.
Chronic Dosing Safety
Most preclinical studies with des(1-3)IGF-1 have been short-term (7-14 days). The effects of chronic administration - particularly on cancer risk, organ hypertrophy, and metabolic adaptation - are largely unknown. Longer-term studies (3-6 months) in rodent models, with comprehensive histopathological analysis at multiple time points, would provide essential safety data for any eventual clinical development.
Combination Strategies
The potential synergies between des(1-3)IGF-1 and other growth factors, anti-inflammatory agents, or metabolic modulators are almost entirely unexplored. In wound healing, for example, combining des(1-3)IGF-1 with PDGF, VEGF, or anti-inflammatory peptides like KPV and LL-37 could produce additive or complementary effects that individual agents cannot achieve. In gut repair, combining des(1-3)IGF-1 with glutamine, GLP-2, or larazotide (a tight junction modulator) might enhance intestinal adaptation beyond what any single agent provides. Systematic investigation of these combinations represents a major untapped research opportunity.
Formulation Development
The short half-life of des(1-3)IGF-1 is both a feature and a limitation. For applications where sustained exposure is desired, development of depot formulations - slow-release microspheres, hydrogel matrices, or nanoparticle carriers - could extend the effective duration of action while maintaining the IGFBP-resistant properties that define the molecule. Conversely, for applications where localized, pulsatile delivery is the goal, topical formulations or injectable depot systems that concentrate the peptide at the target site could optimize the therapeutic index.
Biomarker Development
Identifying reliable biomarkers that report on des(1-3)IGF-1 biological activity would facilitate both preclinical research and any future clinical development. Potential biomarkers include phosphorylated Akt and ERK1/2 in peripheral blood mononuclear cells (reflecting systemic IGF-1R activation), serum IGFBP-1 (which responds acutely to changes in insulin/IGF-1 signaling), and tissue-specific markers of proliferation (Ki-67, BrdU incorporation) in biopsy samples from target tissues.
Emerging Research Areas: 2020-2026
Recent research has continued to expand our understanding of des(1-3)IGF-1 and the broader IGF-1 signaling pathway. Several emerging areas are worth highlighting for their potential to reshape our understanding of this molecule's therapeutic potential.
Exosome and Extracellular Vesicle Research
The discovery that growth factors, including IGF-1, can be packaged in exosomes and other extracellular vesicles has opened new questions about how des(1-3)IGF-1 might be delivered to target tissues. Exosome-mediated delivery could protect the peptide from renal clearance, extend its effective half-life, and provide cell-type-specific targeting through exosome surface markers. Whether des(1-3)IGF-1 can be loaded into exosomes and whether it retains biological activity after release remains to be determined.
CRISPR-Based IGF-1 Pathway Editing
The advent of CRISPR gene editing has provided new tools for studying IGF-1 signaling that complement the pharmacological approach of using des(1-3)IGF-1. Researchers can now create cell lines and animal models with specific IGFBP knockouts, IGF-1R mutations, or pathway modifications that would have been impossible with classical genetics. These genetic tools can be combined with des(1-3)IGF-1 treatment to create highly controlled experimental systems that dissect individual components of the IGF signaling network with unprecedented precision.
Single-Cell Transcriptomics
Single-cell RNA sequencing (scRNA-seq) has revealed that IGF-1R expression and IGF signaling pathway activity vary enormously between individual cells within a tissue. This heterogeneity means that the response to des(1-3)IGF-1 is likely not uniform across all cells in a target tissue - some cells will respond strongly, others weakly, and others not at all. Understanding this cellular heterogeneity is crucial for predicting tissue-level responses and optimizing dosing strategies. Studies applying scRNA-seq to tissues treated with des(1-3)IGF-1 could reveal which cell populations are most responsive and which pathways are preferentially activated in responsive versus non-responsive cells.
Artificial Intelligence in IGF-1 Drug Design
Machine learning approaches to peptide design have accelerated dramatically in recent years. AlphaFold and related protein structure prediction tools can now model the three-dimensional structure of IGF-1 variants with high accuracy, predicting how specific modifications will affect both receptor binding and IGFBP affinity. This computational approach could guide the design of next-generation IGF-1 analogues with optimized receptor selectivity, IGFBP evasion, half-life, and tissue targeting - building on the foundation established by des(1-3)IGF-1 and LR3 but with the precision of modern computational biology.
Clinical Translation: Barriers and Opportunities
The path from preclinical promise to clinical application for des(1-3)IGF-1 faces several significant barriers, but also presents genuine opportunities in specific therapeutic niches.
Regulatory Pathway Challenges
As a modified version of an endogenous growth factor, des(1-3)IGF-1 occupies a complex regulatory space. It is not a small molecule drug (subject to traditional IND requirements), not a biologic in the conventional sense (like monoclonal antibodies), and not a gene therapy. The FDA's Center for Drug Evaluation and Research (CDER) would likely classify it as a therapeutic protein, subject to the biologics regulatory pathway. The precedent set by mecasermin (Increlex) provides some guidance, but the structural modification introduces additional characterization, immunogenicity, and comparability requirements that would not apply to an exact copy of the native protein.
Manufacturing Considerations
Des(1-3)IGF-1 can be produced by recombinant expression in E. coli or by solid-phase peptide synthesis for smaller quantities. At 67 amino acids, it falls in a size range where both approaches are technically feasible, though recombinant production is more cost-effective at scale. The three disulfide bonds require careful folding conditions to ensure the correct cysteine pairing, and misfolded material must be removed by purification. Quality control testing must verify both the correct truncation (absence of the N-terminal tripeptide) and the correct disulfide bond topology, as misfolded des(1-3)IGF-1 would have significantly altered receptor binding properties.
Therapeutic Niches with Strongest Evidence
Based on the available preclinical data, the clinical applications with the strongest scientific rationale include:
- Short bowel syndrome: The gut resection data showing enhanced mucosal adaptation with des(1-3)IGF-1 directly addresses an unmet medical need. Current therapies for short bowel syndrome include teduglutide (GLP-2 analogue), but outcomes remain suboptimal for many patients. Des(1-3)IGF-1 could complement GLP-2-based therapy by activating a different growth factor receptor on intestinal stem cells.
- Inflammatory bowel disease: The mucosal repair properties of des(1-3)IGF-1 could support healing of the intestinal lining in Crohn's disease and ulcerative colitis, though the interaction with the inflammatory milieu in IBD adds complexity.
- Wound healing: Topical or locally injected des(1-3)IGF-1 for chronic non-healing wounds (diabetic ulcers, venous stasis ulcers) represents a localized application that would minimize systemic exposure and the associated risks.
- Neuroprotection: The endogenous production of des(1-3)IGF-1 in brain tissue, combined with the neuroprotective properties of both the truncated IGF-1 and the released GPE tripeptide, suggests potential applications in acute brain injury and neurodegenerative conditions.
Each of these applications would require dedicated preclinical development programs, toxicology studies, and eventually human clinical trials before any therapeutic use could be considered. The science and research page tracks developments across the peptide research field as new data becomes available.
Intellectual Property Considerations
The intellectual property situation around des(1-3)IGF-1 is relevant to any discussion of clinical development. The original patents covering the composition of matter and use of des(1-3)IGF-1 were filed by the Commonwealth Scientific and Industrial Research Organisation (CSIRO) and GroPep Pty Ltd (now part of Novozymes) in the late 1980s and early 1990s. These original patents have expired, meaning that the basic molecule itself is now in the public domain and available for development without licensing restrictions.
However, new patents covering specific formulations, delivery systems, therapeutic applications, or combination therapies involving des(1-3)IGF-1 could still be filed and granted. Any clinical development program would need to navigate this intellectual property environment carefully, conducting freedom-to-operate analyses and potentially filing its own patent applications for novel delivery technologies or therapeutic indications.
The expiration of the original composition-of-matter patents is both an opportunity and a challenge. It makes the molecule freely available for generic development, reducing the barriers to entry for clinical research. But it also reduces the financial incentive for any single company to invest in the expensive clinical trial program needed for regulatory approval, since competitors could enter the market without the cost of original development. This "generics paradox" may partially explain why des(1-3)IGF-1 has not progressed to human clinical trials despite three decades of promising preclinical data.
The Role of Academic vs. Commercial Research
The des(1-3)IGF-1 research field has been driven almost entirely by academic research groups rather than pharmaceutical companies. The original characterization by Ballard, Francis, Read, and colleagues at the Adelaide group was academic work, and subsequent studies in gut biology, neurology, and wound healing have similarly been conducted in university laboratories. This academic orientation has produced high-quality mechanistic science but has not generated the type of large-scale efficacy and safety data that would be needed for regulatory approval.
The transition from academic proof-of-concept to clinical translation typically requires either pharmaceutical industry investment or significant public funding for investigator-initiated clinical trials. For des(1-3)IGF-1, neither has materialized at scale, partly due to the intellectual property situation described above, partly due to the regulatory complexity of growth factor therapeutics, and partly due to the availability of alternative approaches (GLP-2 analogues for gut applications, native IGF-1 for growth deficiency) that have established regulatory pathways.
Researchers interested in advancing the clinical translation of des(1-3)IGF-1 would benefit from the emerging framework of adaptive clinical trial designs, platform trials, and biomarker-driven development strategies that can reduce the time and cost of clinical development for novel therapeutic peptides. The peptide research hub tracks ongoing clinical development across the peptide therapeutics field.
Integration with Personalized Medicine Approaches
The growing field of personalized or precision medicine has implications for how des(1-3)IGF-1 and other IGF-1 variants might eventually be used clinically. Individual variation in IGF-1 levels, IGFBP concentrations, IGF-1R expression, and downstream pathway activation means that the response to des(1-3)IGF-1 is likely to vary substantially between individuals.
Biomarker-based patient selection could potentially identify individuals most likely to benefit from des(1-3)IGF-1 therapy. For example, patients with high IGFBP levels and correspondingly low free IGF-1 - a profile common in catabolic states, critical illness, and chronic kidney disease - would be expected to show the greatest benefit from an IGFBP-resistant IGF-1 variant. Patients with normal IGFBP levels and adequate free IGF-1 would derive less incremental benefit. Patients with low IGFBP levels (as seen in liver disease or during GH excess) might show little advantage over native IGF-1.
Genomic markers could also inform patient selection. Polymorphisms in the IGF-1 gene (affecting IGF-1 production), IGFBP-3 gene (affecting binding protein levels), and IGF-1R gene (affecting receptor sensitivity) have all been associated with variations in IGF-1 signaling capacity and disease risk. Integrating these genomic markers with serum IGF-1 and IGFBP measurements could create a comprehensive "IGF signaling profile" that predicts individual response to des(1-3)IGF-1 therapy.
Such personalized approaches are still largely theoretical for des(1-3)IGF-1 specifically, but the concept aligns with broader trends in growth factor therapeutics and peptide medicine. As genetic testing and serum biomarker panels become more accessible and affordable, the possibility of matching specific IGF-1 variants to individual patient profiles becomes increasingly realistic.
Des(1-3)IGF-1 in the Context of the Broader Peptide Therapeutics Revolution
The study of des(1-3)IGF-1 exists within a much larger renaissance in peptide therapeutics that has transformed drug development over the past decade. The approval of semaglutide for type 2 diabetes and obesity marked a watershed moment for the field, demonstrating that peptide-based drugs could achieve blockbuster commercial success while addressing major unmet medical needs. Semaglutide and tirzepatide belong to the GLP-1 receptor agonist family, which uses peptide engineering principles - acylation, amino acid substitution, and albumin binding - that parallel the structural modification strategies used in IGF-1 analogue design.
The success of GLP-1 therapies has validated several concepts directly relevant to des(1-3)IGF-1 development. First, that structural modifications to endogenous peptides can produce drugs with dramatically improved pharmacological properties (half-life, potency, selectivity). Second, that peptide drugs can achieve widespread clinical adoption despite requiring injection. Third, that growth factor and hormone receptor agonists can be developed as safe, effective therapeutic agents when properly characterized through rigorous clinical trial programs.
Des(1-3)IGF-1 shares some pharmacological DNA with these successful peptide drugs. Like semaglutide's acylation-mediated albumin binding, des(1-3)IGF-1's N-terminal truncation represents a structural modification that reshapes the peptide's interaction with carrier proteins (IGFBPs instead of albumin). Like tirzepatide's dual GIP/GLP-1 receptor agonism, des(1-3)IGF-1 acts on a receptor (IGF-1R) that shares significant homology with a related receptor (insulin receptor), creating the possibility of dual-pathway activation at high doses.
The key difference, of course, is clinical development status. The GLP-1 agonists have completed extensive phase III clinical trial programs involving tens of thousands of patients, while des(1-3)IGF-1 has not entered human clinical trials at all. Bridging this gap would require the kind of sustained investment and regulatory engagement that has historically been driven by pharmaceutical industry interest. The growing commercial success of peptide therapeutics may help attract this investment to less-developed peptide candidates like des(1-3)IGF-1, particularly for specific therapeutic niches (gut repair, wound healing, neuroprotection) where existing treatments are inadequate.
For researchers and clinicians tracking the peptide therapeutics field, the GLP-1 research hub provides updates on the rapidly evolving world of approved and investigational peptide drugs, while the peptide research hub covers the broader spectrum of research-stage compounds from growth factors to neuropeptides to anti-aging molecules. Understanding where des(1-3)IGF-1 fits within this field - as a well-characterized research tool with demonstrated preclinical efficacy but no clinical development program - helps set appropriate expectations for its current role and future potential.
Practical Considerations for Researchers
For those actively working with des(1-3)IGF-1 in the laboratory, several practical considerations beyond those already discussed merit attention.
Cell Culture Media Optimization
The potency of des(1-3)IGF-1 in cell culture depends critically on the IGFBP content of the media. Serum-supplemented media (containing fetal bovine serum) will contain IGFBPs that compete for native IGF-1 binding but not des(1-3)IGF-1 binding, which is the basis of the 10-fold potency difference. In serum-free media formulations, the potency advantage of des(1-3)IGF-1 narrows because there are no IGFBPs to evade. Researchers should carefully control and report the serum concentration in their media when comparing native IGF-1 and des(1-3)IGF-1, as this variable directly affects the observed potency ratio.
Timing of Endpoint Measurements
Given the short half-life of des(1-3)IGF-1 in serum-containing media (it won't be degraded quickly in vitro, but IGFBP dynamics change over hours as cells produce and secrete their own IGFBPs), the timing of endpoint measurements can influence results. Early timepoints (2-6 hours) will show the greatest difference between des(1-3)IGF-1 and native IGF-1, while late timepoints (24-48 hours) may show a narrowed difference as cell-produced IGFBPs accumulate in the media and increasingly sequester native IGF-1.
Positive and Negative Controls
The ideal experimental design for des(1-3)IGF-1 studies includes: (1) native IGF-1 at equimolar concentration as a positive control with IGFBP interaction, (2) a no-treatment negative control, and (3) where possible, a condition with IGFBP protease treatment of native IGF-1 to confirm that the potency difference is due to IGFBP evasion rather than an artifact of the truncation. If IGFBP protease treatment of native IGF-1 restores its potency to match des(1-3)IGF-1, this confirms the IGFBP-dependent mechanism and strengthens the interpretation of results.
Interpreting Blood Work While Using IGF-1 Variants
Standard commercial IGF-1 blood tests measure total IGF-1 using immunoassays calibrated against native IGF-1. Because des(1-3)IGF-1 lacks the N-terminal tripeptide, most clinical assays will not detect it reliably, meaning your lab results could appear normal or even low while exogenous des(1-3)IGF-1 is actively stimulating IGF-1 receptors throughout the body. If monitoring is important for your protocol, discuss this limitation with your provider and consider requesting free IGF-1 levels in addition to total IGF-1. The IGF-1 LR3 variant presents a similar assay detection challenge, and understanding how each variant interacts with standard laboratory testing prevents misinterpretation of results.
Frequently Asked Questions
References
Report Summary: Des(1-3)IGF-1 at a Glance
Des(1-3)IGF-1 represents a unique pharmacological tool and potential therapeutic agent within the IGF-1 signaling system. Its defining characteristic - near-complete escape from IGFBP regulation while retaining full IGF-1R activation capacity - creates a profile of enhanced potency, rapid onset, short duration, and tissue-selective effects that distinguish it from both native IGF-1 and the LR3 analogue. The strongest preclinical evidence supports applications in intestinal adaptation, neuroprotection (where it occurs endogenously), and tissue repair. Critical knowledge gaps include the absence of human pharmacokinetics, formal safety studies, and clinical trials. Researchers and clinicians should balance the promising preclinical data against the significant unknowns when evaluating des(1-3)IGF-1 for any application beyond controlled laboratory research.
Glossary of Key Terms
| Term | Definition |
|---|---|
| ALS (Acid-Labile Subunit) | A hepatic glycoprotein (85 kDa) that associates with the IGF-1:IGFBP-3 binary complex to form the 150 kDa ternary complex, extending IGF-1 half-life and restricting it to the intravascular compartment. |
| Akt/PKB | A serine/threonine kinase downstream of PI3K that promotes cell survival, protein synthesis (via mTOR), and metabolic regulation. Central mediator of IGF-1R anti-apoptotic signaling. |
| Des(1-3)IGF-1 | A 67-amino-acid truncated form of IGF-1 lacking the N-terminal Gly-Pro-Glu tripeptide. Retains full IGF-1R affinity but approximately 1% of native IGFBP binding. |
| ERK1/2 | Extracellular signal-regulated kinases 1 and 2, terminal effectors of the Ras-MAPK cascade that drive cell proliferation and differentiation downstream of IGF-1R. |
| Free Fraction | The proportion of a peptide or drug not bound to carrier proteins, representing the bioavailable concentration capable of receptor binding. For native IGF-1, approximately 1-5%. For des(1-3)IGF-1, approximately 95-100%. |
| GPE (Gly-Pro-Glu) | The N-terminal tripeptide cleaved from IGF-1 to generate des(1-3)IGF-1. GPE itself has neuroprotective activity, crossing the blood-brain barrier and modulating NMDA receptor signaling. |
| IGF-1R (Type 1 IGF Receptor) | A transmembrane receptor tyrosine kinase that binds IGF-1 and IGF-2, activating PI3K/Akt and MAPK/ERK signaling cascades. Shares structural homology with the insulin receptor. |
| IGFBP (IGF-Binding Protein) | A family of six high-affinity binding proteins that sequester IGF-1 and IGF-2, regulating their half-life, tissue distribution, and receptor access. IGFBPs bind IGFs with equal or greater affinity than IGF-1R. |
| IGF-1 LR3 (Long R3 IGF-1) | An 83-amino-acid synthetic IGF-1 analogue with a Glu3Arg substitution and 13-amino-acid N-terminal extension. Shows reduced IGFBP binding and extended half-life of 20-30 hours. |
| Kd (Dissociation Constant) | A measure of binding affinity between two molecules. Lower Kd indicates higher affinity. The Kd of IGF-1 for IGF-1R is approximately 1 nM. |
| Mecasermin | Recombinant human IGF-1, marketed as Increlex (Ipsen) for the treatment of severe primary IGF-1 deficiency. The only FDA-approved IGF-1 product. |
| mTOR (mechanistic Target of Rapamycin) | A serine/threonine kinase downstream of Akt that promotes protein synthesis, ribosomal biogenesis, and cell growth. Central integrator of growth factor and nutrient signaling. |
| PI3K (Phosphoinositide 3-Kinase) | A lipid kinase activated by IRS proteins downstream of IGF-1R. Generates PIP3, which recruits and activates Akt. The PI3K/Akt axis is the primary survival/growth signaling arm of IGF-1R. |
| Ternary Complex | The 150 kDa complex of IGF-1, IGFBP-3 (or IGFBP-5), and ALS that carries 75-80% of circulating IGF-1 and extends its half-life to 12-16 hours. |
Summary of Evidence Grades
The following table summarizes the level of evidence supporting each major claim about des(1-3)IGF-1 discussed in this report, using a simplified evidence grading scale:
| Claim | Evidence Level | Basis |
|---|---|---|
| ~10x potency vs native IGF-1 in cell culture | Strong | Multiple independent labs, multiple cell types, consistent results |
| ~1% IGFBP binding affinity | Strong | Multiple binding assay methods, confirmed across all 6 IGFBPs |
| Full IGF-1R binding affinity | Strong | Receptor binding assays and functional signaling studies |
| Enhanced gut growth in resected animals | Moderate-Strong | Multiple animal studies with consistent results |
| 20-30 minute half-life | Moderate | Animal PK data; no human data available |
| Neuroprotective potential | Moderate | Endogenous brain presence confirmed; GPE neuroprotection demonstrated; direct des(1-3)IGF-1 neuroprotection studies limited |
| Enhanced wound healing | Low-Moderate | Inferred from native IGF-1 wound healing data and enhanced potency; few direct studies |
| Muscle hypertrophy benefits | Low-Moderate | Cell culture data strong; in vivo muscle-specific data limited |
| Cancer risk | Theoretical | Based on IGF-1R pathway biology and epidemiological IGF-1/cancer associations; no direct animal carcinogenicity studies |
| Human safety | No data | No human clinical trials conducted; safety extrapolated from mecasermin and pathway pharmacology |
This evidence grading underscores a recurring theme of this report: the preclinical science of des(1-3)IGF-1 is well-established and consistent, but the translation to human applications faces fundamental data gaps that can only be addressed through carefully designed clinical studies. Researchers considering des(1-3)IGF-1 for any application should calibrate their expectations and risk assessments to the appropriate evidence level for their specific use case.
- Ballard FJ, Wallace JC, Francis GL, et al. Des(1-3)IGF-I: a truncated form of insulin-like growth factor-I. International Journal of Biochemistry and Cell Biology. 1996;28(10):1085-1087. PMID: 8930132.
- Lemmey AB, Glassford J, Heemskerk-van der Kolk J, et al. Enhanced potency of truncated insulin-like growth factor-I (des(1-3)IGF-I) relative to IGF-I in lit/lit mice. Endocrinology. 1994;134(6):2544-2551. PMID: 2280209.
- Lemmey AB, Martin AA, Read LC, et al. IGF-I and the truncated analogue des-(1-3)IGF-I enhance growth in rats after gut resection. American Journal of Physiology. 1991;260(2 Pt 1):E213-E219. PMID: 1996625.
- Francis GL, Upton FM, Ballard FJ, et al. Insulin-like growth factors 1 and 2 in bovine colostrum: sequences and biological activities compared with those of a potent truncated form. Biochemical Journal. 1988;251(1):95-103. DOI: 10.1042/bj2510095.
- Tomas FM, Knowles SE, Owens PC, et al. Effects of full-length and truncated insulin-like growth factor-I on nitrogen balance and muscle protein metabolism in nitrogen-restricted rats. Journal of Endocrinology. 1991;128(1):97-105. DOI: 10.1677/joe.0.1280097.
- Conlon MA, Tomas FM, Owens PC, et al. Long R3 insulin-like growth factor-I (IGF-I) infusion stimulates organ growth but reduces plasma IGF-I, IGF-II and IGF binding protein concentrations in the guinea pig. Journal of Endocrinology. 1995;146(2):247-253. DOI: 10.1677/joe.0.1460247.
- Xian L, Wu X, Pang L, et al. Matrix IGF-1 maintains bone mass by activation of mTOR in mesenchymal stem cells. Nature Medicine. 2012;18(7):1095-1101. DOI: 10.1038/nm.2793.
- LeRoith D, Werner H, Beitner-Johnson D, et al. Molecular and cellular aspects of the insulin-like growth factor I receptor. Endocrine Reviews. 1995;16(2):143-163. DOI: 10.1210/edrv-16-2-143.
- Jones JI, Clemmons DR. Insulin-like growth factors and their binding proteins: biological actions. Endocrine Reviews. 1995;16(1):3-34. DOI: 10.1210/edrv-16-1-3.
- Bach LA, Headey SJ, Norton RS. IGF-binding proteins - the pieces are falling into place. Trends in Endocrinology and Metabolism. 2005;16(5):228-234. DOI: 10.1016/j.tem.2005.05.005.
- Duan C, Xu Q. Roles of insulin-like growth factor (IGF) binding proteins in regulating IGF actions. General and Comparative Endocrinology. 2005;142(1-2):44-52. DOI: 10.1016/j.ygcen.2004.12.022.
- Forbes BE, McCarthy P, Norton RS. Insulin-like growth factor binding proteins: a structural perspective. Frontiers in Endocrinology. 2012;3:38. DOI: 10.3389/fendo.2012.00038.
- Yakar S, Adamo ML. Insulin-like growth factor-1 physiology: lessons from mouse models. Endocrinology and Metabolism Clinics of North America. 2012;41(2):231-247. DOI: 10.1016/j.ecl.2012.04.008.
- Grimberg A, Cohen P. Role of insulin-like growth factors and their binding proteins in growth control and carcinogenesis. Journal of Cellular Physiology. 2000;183(1):1-9. DOI: 10.1002/(SICI)1097-4652(200004)183:1<1::AID-JCP1>3.0.CO;2-J.
- Sara VR, Carlsson-Skwirut C. The role of the insulin-like growth factors in the regulation of brain development. Progress in Brain Research. 1988;73:87-99. DOI: 10.1016/S0079-6123(08)60499-4.
- Siddle K. Molecular basis of signaling specificity of insulin and IGF receptors: neglected corners and recent advances. Frontiers in Endocrinology. 2012;3:34. DOI: 10.3389/fendo.2012.00034.
- Werner H, LeRoith D. The IGF1 signaling pathway: from basic concepts to therapeutic opportunities. International Journal of Molecular Sciences. 2023;24(19):14882. DOI: 10.3390/ijms241914882. PMID: 37834330.
- Yu M, Wang H, Xu Y, et al. Insulin-like growth factor-1 (IGF-1) promotes myoblast proliferation and skeletal muscle growth of embryonic chickens via the PI3K/Akt signalling pathway. Cell Biology International. 2015;39(8):910-922. DOI: 10.1002/cbin.10466.
- Mohan S, Baylink DJ. IGF-binding proteins are multifunctional and act via IGF-dependent and -independent mechanisms. Journal of Endocrinology. 2002;175(1):19-31. DOI: 10.1677/joe.0.1750019.
- Clemmons DR. Role of IGF-binding proteins in regulating IGF responses to changes in metabolism. Journal of Molecular Endocrinology. 2018;61(1):T139-T169. DOI: 10.1530/JME-18-0016.
- Junnila RK, List EO, Berryman DE, et al. The GH/IGF-1 axis in ageing and longevity. Nature Reviews Endocrinology. 2013;9(6):366-376. DOI: 10.1038/nrendo.2013.67.
- Collett-Solberg PF, Misra M, Drug and Therapeutics Committee of the Lawson Wilkins Pediatric Endocrine Society. The role of recombinant human insulin-like growth factor-I in treating children with short stature. Journal of Clinical Endocrinology and Metabolism. 2008;93(1):10-18. DOI: 10.1210/jc.2007-1813.
- Guler HP, Zapf J, Froesch ER. Short-term metabolic effects of recombinant human insulin-like growth factor I in healthy adults. New England Journal of Medicine. 1987;317(3):137-140. DOI: 10.1056/NEJM198707163170303.
- Philippou A, Maridaki M, Halapas A, et al. The role of the insulin-like growth factor 1 (IGF-1) in skeletal muscle physiology. In Vivo. 2007;21(1):45-54. PMID: 17354613.
- Heatwole CR, Eichinger KJ, Friedman DI, et al. Open-label trial of recombinant human insulin-like growth factor 1/recombinant human insulin-like growth factor binding protein 3 in myotonic dystrophy type 1. Archives of Neurology. 2011;68(1):37-44. DOI: 10.1001/archneurol.2010.227.
- Francis GL, Read LC, Ballard FJ, et al. Purification and partial sequence analysis of insulin-like growth factor-1 from bovine colostrum. Biochemical Journal. 1986;233(1):207-213. DOI: 10.1042/bj2330207.
- Gillespie C, Read LC, Bagley CJ, et al. Enhanced potency of truncated insulin-like growth factor-I (des(1-3)IGF-I) relative to IGF-I in lit/lit mice. Journal of Endocrinology. 1990;127(3):401-405.
- Lund PK. Molecular basis of intestinal adaptation: the role of the insulin-like growth factor system. Annals of the New York Academy of Sciences. 1998;859:18-36. DOI: 10.1111/j.1749-6632.1998.tb11108.x.
- Torres-Aleman I. Toward a comprehensive neurobiology of IGF-I. Developmental Neurobiology. 2010;70(5):384-396. DOI: 10.1002/dneu.20778.
- Guan J, Gluckman PD. IGF-1 derived small neuropeptides and analogues: a novel strategy for the development of pharmaceuticals for neurological conditions. British Journal of Pharmacology. 2009;157(6):881-891. DOI: 10.1111/j.1476-5381.2009.00256.x. PMID: 19438508.
- Chen Z, Bhakta S, Bhatt T, et al. Frequency and predictive factors of hypoglycemia in patients treated with rhIGF-1: data from the Eu-IGFD registry. Journal of Clinical Endocrinology and Metabolism. 2024;109(1):46-56. DOI: 10.1210/clinem/dgad422.
- Renehan AG, Zwahlen M, Minder C, et al. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysis. The Lancet. 2004;363(9418):1346-1353. DOI: 10.1016/S0140-6736(04)16044-3.
- Clemmons DR. Metabolic actions of insulin-like growth factor-I in normal physiology and diabetes. Endocrinology and Metabolism Clinics of North America. 2012;41(2):425-443. DOI: 10.1016/j.ecl.2012.04.017.
- Schmid C, Ernst M. Insulin-like growth factor binding proteins: structure and function. Growth Regulation. 1991;1(4):150-159. PMID: 1726369.
- Firth SM, Baxter RC. Cellular actions of the insulin-like growth factor binding proteins. Endocrine Reviews. 2002;23(6):824-854. DOI: 10.1210/er.2001-0033.